
Содержание
сообщение на тему «простейшие» — Школьные Знания.com
Простейшие — это одноклеточные разнообразные животные,которые обитают в водоемах и во влажной почве.Простейшие способны жить самостоятельно.
Простейших делят на три типа:
• Саркомагистофоры
• Споровики
• Инфузории
Тип Саркомагистрофоры включает в себя саркодовые и жгутиковые.
Тип Инфузорий включает в себя ресничные и сосущие.
Давайте расскажем о строении,дыхании,выделении,размножении и питании одноклеточных (или простейших) животных.
•Строение•
Для простейших характерно разделение цитоплазмы на два слоя: внутренний (эндоплазма) и внешний (наружный или эктоплазма).
Строение внутреннего слоя включает в себя органеллы и ядро.
Особенностью строения простейших является наличие структур,принадлежавшим только им. Например,порошицы,сократительная вакуоль,клеточная глотка и клеточный рот.
Например,порошицы,сократительная вакуоль,клеточная глотка и клеточный рот.
•Дыхание•
Способы дыхания у простейших зависит от среды обитания.
Свободноживущие одноклеточные используют кислород.
Паразитические одноклеточные обитают,как известно,в бескилородной среде,дыхание которого происходит при помощи гликолиза.
Гликолиз — это расщепление углеводов в бескилородной среде.
•Выделение•
Выделение у простеших осуществляется через сократительные вакуоли или диффузию.
•Размножение•
У простейших два способа размножения: половое и бесполое.
Бесполое размножение представлено митозом,во время которого происходит деление ядра,а затем — цитоплазмы.
Половое размножение происходит при помощи оогамии,изогамии и анизогамии.
•Питание•
У простейших также возможно два типа питания: гетеротрофный и смешанный.
Различают два способа пищи: фагоцитоз и пиноцитоз.
Фаготицоз — процесс захвата твердых частиц пищи при помощи вырост цитоплазмы.
Пиноцитоз — процесс захвата твердых частиц пищи самой клеточной поверхностью.
==================================================================
Простейшие (одноклеточные организмы) | Микробиология. Реферат, доклад, сообщение, кратко, презентация, лекция, шпаргалка, конспект, ГДЗ, тест
Царство животных организмов является одним из самых крупных подразделений органического мира. Животные возникли около 1-1,5 млрд лет назад в морской среде в форме клеток, напоминающих микроскопических бесхлорофильных амебоидных жгутиконосцев.
Большой интерес представляют эукариотические организмы, обладающие признаками животных и находящиеся на клеточном уровне организации, — простейшие (рис. 95).
Простейшие — это одноклеточные организмы. Подцарство простейших — разнообразная и процветающая группа животных, состоящая примерно из 70 тыс. современных видов. Традиционно изучение животных начинают с простейших, являющих собой исходный этап в эволюции жизни.
Тело простейших морфологически представлено одной клеткой, а функционально соответствует целому организму. Среди них есть организмы, которые на свету питаются как растения, а в темноте — как животные (например, эвглена зелёная). Именно в этой группе организмов проходит «граница» между растительным и животным миром.
| Рис. 95. Простейшие: а — амёба; б — инфузория туфелька; в — эвглена зелёная |
Как и прокариоты, простейшие легко приспосабливаются к самым экстремальным условиям изменяющейся среды, а потому присутствуют во всех частях биосферы.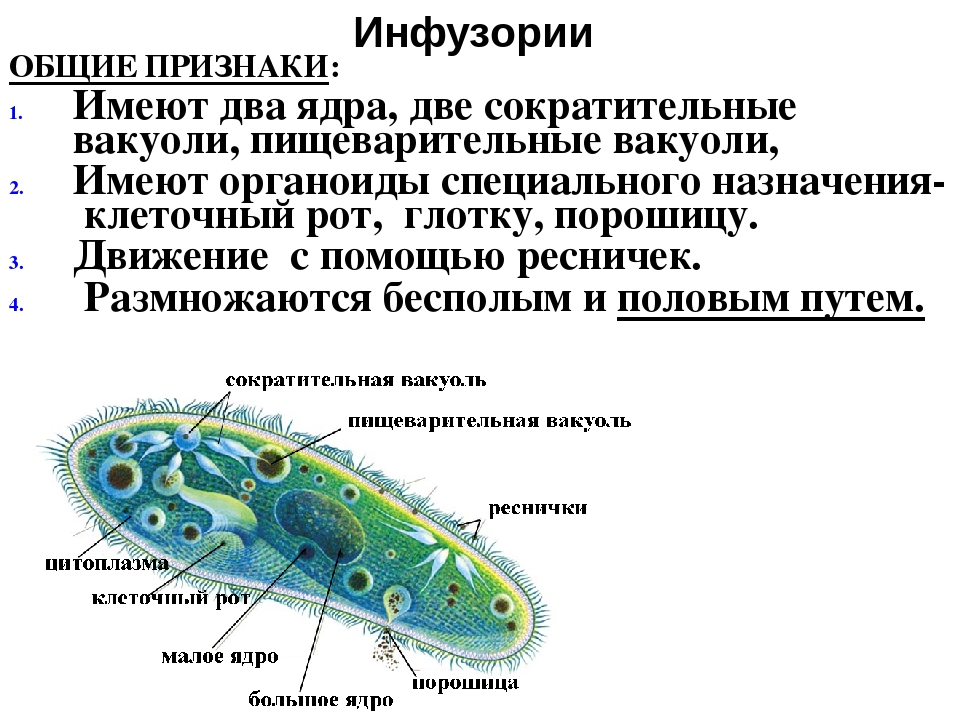 Они могут переходить в состояние покоя, если условия становятся неблагоприятными для жизни, и делаются активными, если обстановка улучшается. Делятся простейшие очень интенсивно, приблизительно раз в 3 часа. Материал с сайта http://doklad-referat.ru
Они могут переходить в состояние покоя, если условия становятся неблагоприятными для жизни, и делаются активными, если обстановка улучшается. Делятся простейшие очень интенсивно, приблизительно раз в 3 часа. Материал с сайта http://doklad-referat.ru
Размер самых маленьких из них — 2-4 мкм (1 мкм = 0,001 мм), наиболее крупных — до 1000 мкм (1 мм). Лишь ископаемые раковинные корненожки (нуммулиты) достигали нескольких сантиметров. Поэтому изучают простейших с помощью оптического микроскопа, увеличивающего изображение объекта в 2,5-3 тыс. раз. Многие структуры можно увидеть лишь с помощью электронного микроскопа, увеличивающего изображение в 200-300 тыс. раз, что позволяет рассмотреть даже крупные молекулы клеток живых организмов.
Роль одноклеточных животных зависит от места обитания, особенностей биологии, питания и численности.
На этой странице материал по темам:
Презентация на тему заболевания вызванные простейшими
Простейшие краткий доклад
Многоклеточные организмы доклад
Положительная и отрицательная роль одноклеточных животных
Какое простое вызывает малярию по биологии
Вопросы по этому материалу:
Дайте характеристику типа Простейшие.

Охарактеризуйте простейших с точки зрения их строения и функционирования.
Назовите представителей типа Простейшие.
Простейшие [Одноклеточные] — животное, описание, характеристика, строение, питание, дыхание, размножение, где обитает, виды, фото, вики — WikiWhat
История открытия
Как известно, в 1675 г. т. е. более трёхсот лет назад, А. Левенгук открыл «анималькулов» (зверушек), которых впоследствии назвали инфузориями. С 1820 г. установилось название Protozoa, что в переводе с греческого означает «простейшие животные». Зоолог К. Зибольд посчитал их особым типом животного царства и выделил два класса: инфузорий и корненожек. Он же определил, что простота их организации соответствует одной клетке. С тех пор одноклеточность простейших стала общепризнанной, а название «одноклеточные» и «простейшие» стали синонимами.
По уровню организации все живые организмы классифицируются на две группы. Привычное для нас деление на одноклеточных и многоклеточных потребовало уточнения, после того как при изучении строения организмов был применён электронный микроскоп и появились новые методы исследования. Возникли вопросы об основных различиях, определяющих уровни развития, а также о планах строения. Поэтому необходимо рассмотреть организацию простейших — парафилетической группы, объединяющей представителей органического мира, относимых ранее к растениям, животным и грибам, но имеющих свои специфические особенности.
Привычное для нас деление на одноклеточных и многоклеточных потребовало уточнения, после того как при изучении строения организмов был применён электронный микроскоп и появились новые методы исследования. Возникли вопросы об основных различиях, определяющих уровни развития, а также о планах строения. Поэтому необходимо рассмотреть организацию простейших — парафилетической группы, объединяющей представителей органического мира, относимых ранее к растениям, животным и грибам, но имеющих свои специфические особенности.
Происхождение
Самозарождение
Природа простейших долгое время оставалась предметом спора. Одни учёные рассматривали их как живых молекул, или простые комплексы таких молекул, которые способны самозарождаться, т. е. возникать сами по себе. Этих воззрений придерживались немногие учение, тем более что блестящие опыты Л. Спаланцани в XVIII в. Л. Пастера в XIX в. опровергли идею самозарождения.
Целлюляризация
Другие учёные считали простейших весьма сложно организованными существами, которых можно структурно сравнить с высокоорганизованными животными.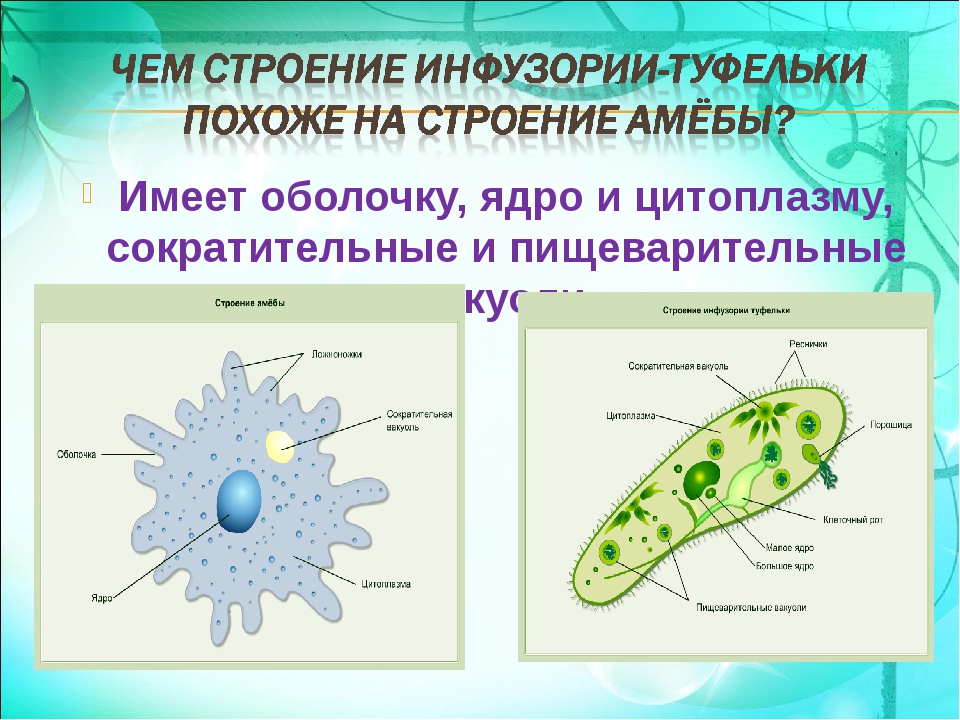 Основание для этого они видели в том, что в организме многоклеточных есть структуры, не имеющие разделения на клетки, например синцитии. Исходя из подобных воззрений, зоолог Й. Хаджи в 50-60-е годы XX в. выдвинул даже теорию происхождения многоклеточных животных путём целлюляризации. Обнаружив сходство инфузорий с самыми примитивными ресничными червями, так называемыми бескишечными, Хаджи предположил, что при обособлении частей тела инфузории, содержащих органоиды, и образовании между ними перегородок возникает многоклеточный организм. Следовательно, по своей природе инфузория сравнима с целым организмом низших многоклеточных. Однако после электронно-микроскопических исследований было доказано, что теория целлюляризации опирается только на внешние аналогии и конвергентные сходства.
Основание для этого они видели в том, что в организме многоклеточных есть структуры, не имеющие разделения на клетки, например синцитии. Исходя из подобных воззрений, зоолог Й. Хаджи в 50-60-е годы XX в. выдвинул даже теорию происхождения многоклеточных животных путём целлюляризации. Обнаружив сходство инфузорий с самыми примитивными ресничными червями, так называемыми бескишечными, Хаджи предположил, что при обособлении частей тела инфузории, содержащих органоиды, и образовании между ними перегородок возникает многоклеточный организм. Следовательно, по своей природе инфузория сравнима с целым организмом низших многоклеточных. Однако после электронно-микроскопических исследований было доказано, что теория целлюляризации опирается только на внешние аналогии и конвергентные сходства.
Клеточная теория Т. Шванна
С позиций клеточной теории, разработанной М. Шлейденом и Т. Шванном, простейшие представляют собой одноклеточные организмы.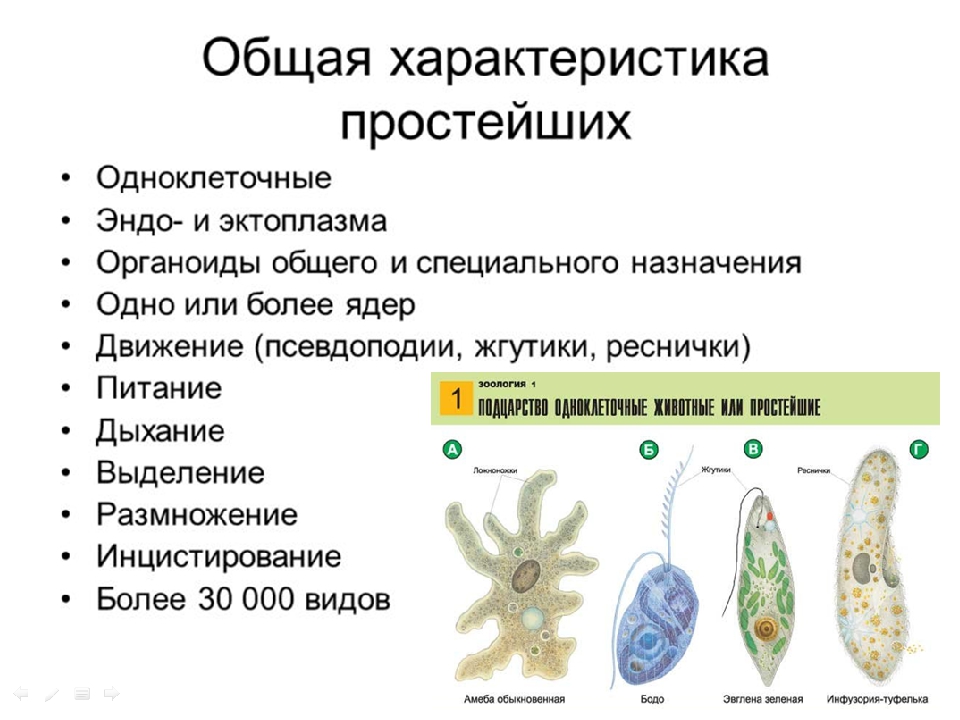 По мнению современных учёных, придерживающихся этих воззрений, простейшие — это клетки, которые функционально являются организмами. Однако функции не могут существовать отдельно от определённых структур. Таким образом, современное определение простейших как микроскопических одноклеточных животных, представляющих собой физиологически самостоятельные организмы, не соответствует нынешнему уровню знаний. Удовлетворительное определение простейших может быть дано после ответов на следующие вопросы: являются ли простейшие только одноклеточными организмами? Всегда ли их размеры микроскопически малы? Являются ли они исключительно животными? Являются ли они организмами только в физиологическом отношении?
По мнению современных учёных, придерживающихся этих воззрений, простейшие — это клетки, которые функционально являются организмами. Однако функции не могут существовать отдельно от определённых структур. Таким образом, современное определение простейших как микроскопических одноклеточных животных, представляющих собой физиологически самостоятельные организмы, не соответствует нынешнему уровню знаний. Удовлетворительное определение простейших может быть дано после ответов на следующие вопросы: являются ли простейшие только одноклеточными организмами? Всегда ли их размеры микроскопически малы? Являются ли они исключительно животными? Являются ли они организмами только в физиологическом отношении?
Размеры простейших варьируют в той же степени, что и размеры многоклеточных. В среднем они колеблются от 5 до 250 мкм. Наиболее мелки внутриклеточные паразиты, например лейшмании (1-4 мкм). Но уже инфузория-туфелька значительно крупнее — 150-250 мкм, амёба протей — 600 мкм, а современные глубоководные радиолярии имеют диаметр тела 2,5 см.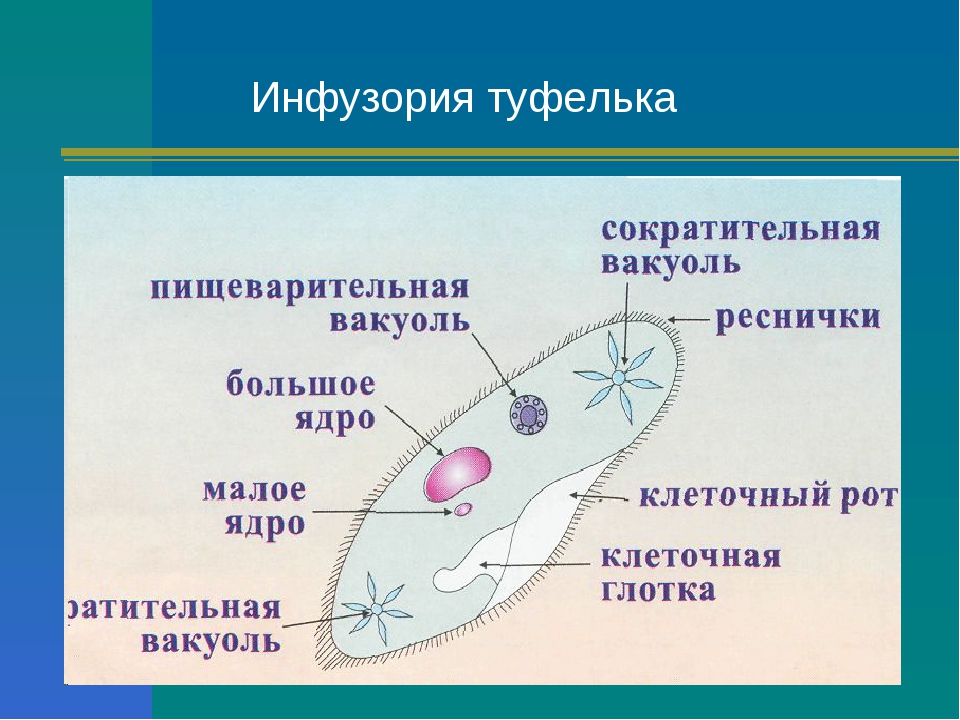 В современную систему простейших входят миксомицеты — настоящие гиганты среди протистов. Длина плазмодия одного из них доходит до 1,2 м. Короче говоря, различия в размерах простейших равняются 106.
В современную систему простейших входят миксомицеты — настоящие гиганты среди протистов. Длина плазмодия одного из них доходит до 1,2 м. Короче говоря, различия в размерах простейших равняются 106.
Раньше считалось, что все простейшие — микроскопические существа. Сегодня среди них обнаружены и макроскопические виды. Размеры простейших колеблются от 1 мк (1 мк = 10-3 мм) до 5 см и более, но в среднем от 5 до 250 мк. Самые мелкие среди них — внутриклеточные паразиты, например некоторые споровики и жгутиконосцы. Самые крупные представители — колониальные радиолярии, размеры которых могут достигать 25 см.
Подцарство Одноклеточные (Простейшие) объединяет животных, тело которых состоит из одной клетки. Она выполняет функции самостоятельного организма. Клетка простейшего состоит из цитоплазмы, органоидов, одного или нескольких ядер. В ней происходят обмен веществ с внешней средой, процессы размножения в развития.
Многие одноклеточные обладают специальными органоидами (движения, питания, выделения), возникшими как результат приспособления к среде обитания.
Клетка — это самовоспроизводящееся образование, отделённое от своего окружения плазматической мембраной, способствующей регуляции обмена между внутренней и внешней средой.
Среда обитания
Простейшие животные — процветающая и разнообразная группа (около 70 000 видов) — обитатели водоёмов и влажной почвы. Преимущественно они входят в состав зоопланктона — совокупность мельчайших животных, обитающих в морских и пресноводных водоёмах. На суше они встречаются тоже в водной среде — в почвенной капельной воде, а также в жидкой среде внутри многоклеточных животных и растений. Хотя почвенные простейшие животные могут существенно влиять на количество бактерий, все же их значение несравненно меньше, чем у простейших в пресных и морских водоёмах.
Жизнедеятельность
см. Питание простейших
Многие простейшие животные так же мелки и просто устроены, как и некоторые клетки крупных животных. Но они отличаются от них тем, что способны жить самостоятельно. Одноклеточные животные представляют собой слаженный организм, осуществляющий питание, дыхание, выделение, размножение, рост, развитие и обмен веществ. У него в протоплазме существует как бы разделение труда: каждая из её обособленных, более мелких образований выполняет свою особую задачу.
Но они отличаются от них тем, что способны жить самостоятельно. Одноклеточные животные представляют собой слаженный организм, осуществляющий питание, дыхание, выделение, размножение, рост, развитие и обмен веществ. У него в протоплазме существует как бы разделение труда: каждая из её обособленных, более мелких образований выполняет свою особую задачу.
Например, ядро регулирует жизнедеятельность всего одноклеточного организма и воспроизводит само себя, благодаря чему образуются новые дочерние организмы; в пищеварительной вакуоли происходит переваривание пищи; сократительная вакуоль удаляет избыток воды и растворенные в ней вредные для организма вещества.
При неблагоприятных условиях многие простейшие перестают питаться, теряют органы движения, покрываются толстой оболочкой и образуют цисту. При наступлении благоприятных условий одноклеточные принимают прежний облик.
Классификация
Согласно названию Protozoa, в это подцарство должны входить только животные. Но в современной системе простейших содержатся зелёные жгутиконосцы (ботаники считают их водорослями), миксомицеты и плазмодиофориды (по мнению микологов, это грибы) и т. д. В связи с этим древние простейшие скорее всего могут рассматриваться в качестве исходной группы, давшей начало и грибам, и растениям, и животным. Поэтому в настоящее время должно считаться признанным выделение особого царства протистов и противопоставление его царствам растений и животных. Выделение царства протистов принадлежит знаменитому зоологу и эволюционисту Э. Геккелю (1866). Protozoa же могут быть выделены в системе протистов в качестве подцарства.
Но в современной системе простейших содержатся зелёные жгутиконосцы (ботаники считают их водорослями), миксомицеты и плазмодиофориды (по мнению микологов, это грибы) и т. д. В связи с этим древние простейшие скорее всего могут рассматриваться в качестве исходной группы, давшей начало и грибам, и растениям, и животным. Поэтому в настоящее время должно считаться признанным выделение особого царства протистов и противопоставление его царствам растений и животных. Выделение царства протистов принадлежит знаменитому зоологу и эволюционисту Э. Геккелю (1866). Protozoa же могут быть выделены в системе протистов в качестве подцарства.
Одноклеточные прошли длительный путь эволюции, в ходе которой возникло их огромное разнообразие. В зависимости от сложности строения и способов передвижения выделяют несколько типов простейших. Материал с сайта http://wikiwhat.ru
Значение и роль
Со времён Линнея и до наших дней простейшие привлекают внимание учёных по разным причинам. Возникла даже специальная наука — протозоология.
Возникла даже специальная наука — протозоология.
Модель клетки
Простейшие представляют особый уровень развития живого мира поэтому их изучение имеет общебиологическое значение. В настоящее время простейшие используются как модель эукариотной клетки для исследований в области молекулярной биологии, генетики, радиобиологии и других наук.
Роль в биосфере
Простейшие играют важную роль в биосфере, являясь необходимым звеном в круговороте веществ и потоке энергии. Простейшие животные — очень важные потребители бактерий и одноклеточных водорослей, а также животных, в основном одноклеточных. По объёму потребляемого органического вещества они занимают третье место вслед за бактериями и грибами. Они образуют начальные звенья в сетях питания после растительных организмов — первичных продуцентов органических веществ.
Возбудители заболеваний
Одноклеточные являются возбудителями заболеваний человека и домашних животных.
Вопросы к этой статье:
Расскажите о разнообразии простейших.
Кто из представителей простейших имеет наиболее сложное строение?
Каково значение простейших в природе и жизни человека?
В чём заключается средообразующая роль простейших?
Простейшие [Одноклеточные] — животное, описание, характеристика, строение, питание, дыхание, размножение, где обитает, виды, фото, вики — WikiWhat
История открытия
Как известно, в 1675 г. т. е. более трёхсот лет назад, А. Левенгук открыл «анималькулов» (зверушек), которых впоследствии назвали инфузориями. С 1820 г. установилось название Protozoa, что в переводе с греческого означает «простейшие животные». Зоолог К. Зибольд посчитал их особым типом животного царства и выделил два класса: инфузорий и корненожек. Он же определил, что простота их организации соответствует одной клетке.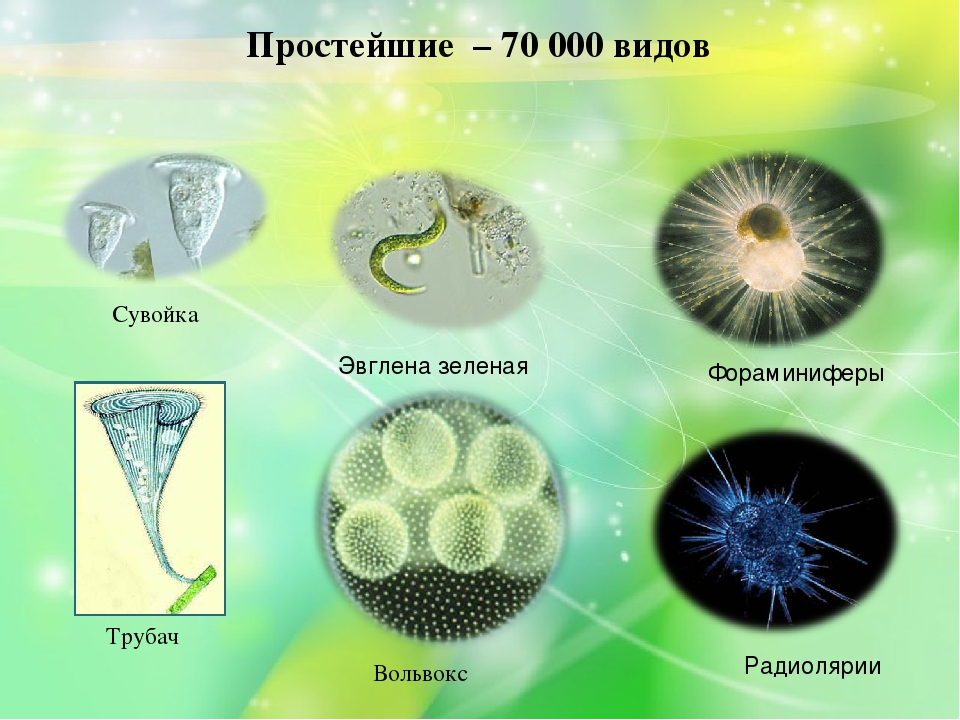 С тех пор одноклеточность простейших стала общепризнанной, а название «одноклеточные» и «простейшие» стали синонимами.
С тех пор одноклеточность простейших стала общепризнанной, а название «одноклеточные» и «простейшие» стали синонимами.
По уровню организации все живые организмы классифицируются на две группы. Привычное для нас деление на одноклеточных и многоклеточных потребовало уточнения, после того как при изучении строения организмов был применён электронный микроскоп и появились новые методы исследования. Возникли вопросы об основных различиях, определяющих уровни развития, а также о планах строения. Поэтому необходимо рассмотреть организацию простейших — парафилетической группы, объединяющей представителей органического мира, относимых ранее к растениям, животным и грибам, но имеющих свои специфические особенности.
Происхождение
Самозарождение
Природа простейших долгое время оставалась предметом спора. Одни учёные рассматривали их как живых молекул, или простые комплексы таких молекул, которые способны самозарождаться, т.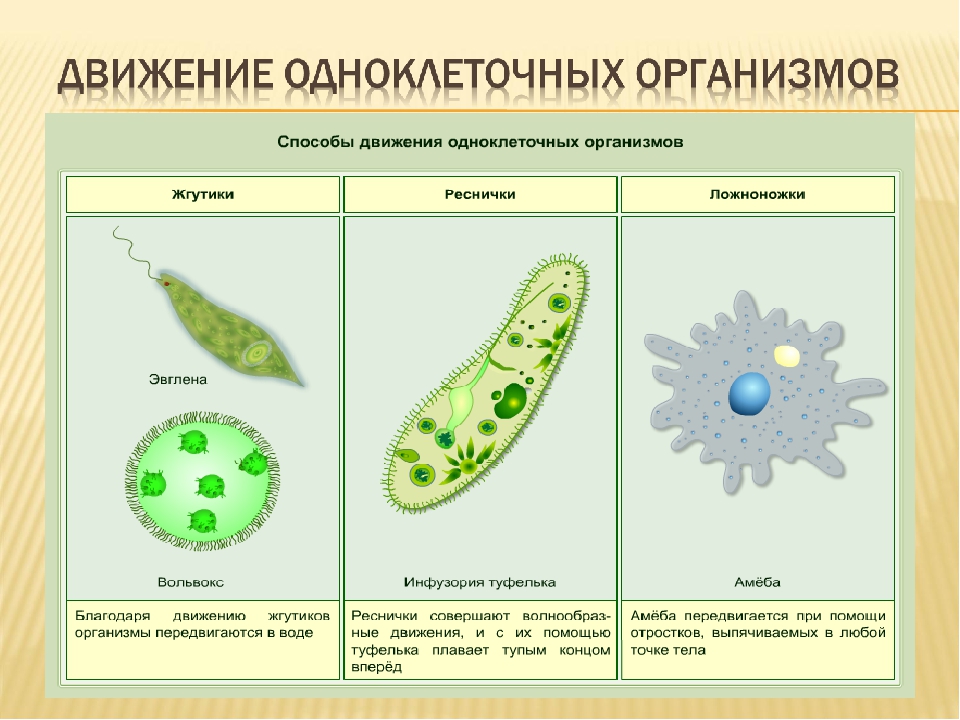 е. возникать сами по себе. Этих воззрений придерживались немногие учение, тем более что блестящие опыты Л. Спаланцани в XVIII в. Л. Пастера в XIX в. опровергли идею самозарождения.
е. возникать сами по себе. Этих воззрений придерживались немногие учение, тем более что блестящие опыты Л. Спаланцани в XVIII в. Л. Пастера в XIX в. опровергли идею самозарождения.
Целлюляризация
Другие учёные считали простейших весьма сложно организованными существами, которых можно структурно сравнить с высокоорганизованными животными. Основание для этого они видели в том, что в организме многоклеточных есть структуры, не имеющие разделения на клетки, например синцитии. Исходя из подобных воззрений, зоолог Й. Хаджи в 50-60-е годы XX в. выдвинул даже теорию происхождения многоклеточных животных путём целлюляризации. Обнаружив сходство инфузорий с самыми примитивными ресничными червями, так называемыми бескишечными, Хаджи предположил, что при обособлении частей тела инфузории, содержащих органоиды, и образовании между ними перегородок возникает многоклеточный организм. Следовательно, по своей природе инфузория сравнима с целым организмом низших многоклеточных.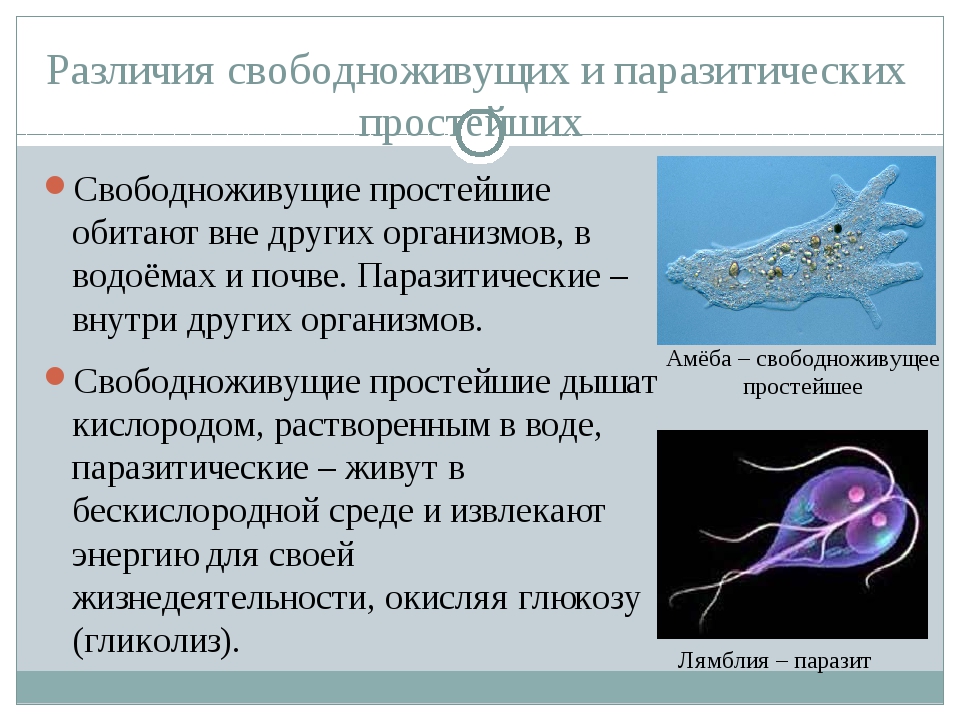 Однако после электронно-микроскопических исследований было доказано, что теория целлюляризации опирается только на внешние аналогии и конвергентные сходства.
Однако после электронно-микроскопических исследований было доказано, что теория целлюляризации опирается только на внешние аналогии и конвергентные сходства.
Клеточная теория Т. Шванна
С позиций клеточной теории, разработанной М. Шлейденом и Т. Шванном, простейшие представляют собой одноклеточные организмы. По мнению современных учёных, придерживающихся этих воззрений, простейшие — это клетки, которые функционально являются организмами. Однако функции не могут существовать отдельно от определённых структур. Таким образом, современное определение простейших как микроскопических одноклеточных животных, представляющих собой физиологически самостоятельные организмы, не соответствует нынешнему уровню знаний. Удовлетворительное определение простейших может быть дано после ответов на следующие вопросы: являются ли простейшие только одноклеточными организмами? Всегда ли их размеры микроскопически малы? Являются ли они исключительно животными? Являются ли они организмами только в физиологическом отношении?
Размеры простейших варьируют в той же степени, что и размеры многоклеточных.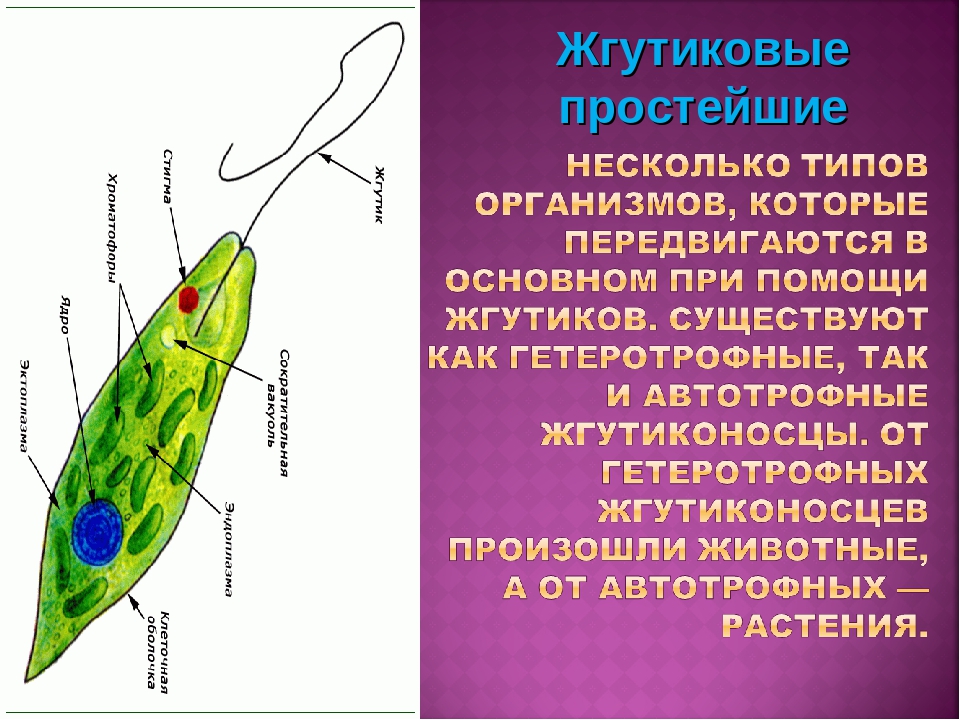 В среднем они колеблются от 5 до 250 мкм. Наиболее мелки внутриклеточные паразиты, например лейшмании (1-4 мкм). Но уже инфузория-туфелька значительно крупнее — 150-250 мкм, амёба протей — 600 мкм, а современные глубоководные радиолярии имеют диаметр тела 2,5 см. В современную систему простейших входят миксомицеты — настоящие гиганты среди протистов. Длина плазмодия одного из них доходит до 1,2 м. Короче говоря, различия в размерах простейших равняются 106.
В среднем они колеблются от 5 до 250 мкм. Наиболее мелки внутриклеточные паразиты, например лейшмании (1-4 мкм). Но уже инфузория-туфелька значительно крупнее — 150-250 мкм, амёба протей — 600 мкм, а современные глубоководные радиолярии имеют диаметр тела 2,5 см. В современную систему простейших входят миксомицеты — настоящие гиганты среди протистов. Длина плазмодия одного из них доходит до 1,2 м. Короче говоря, различия в размерах простейших равняются 106.
Раньше считалось, что все простейшие — микроскопические существа. Сегодня среди них обнаружены и макроскопические виды. Размеры простейших колеблются от 1 мк (1 мк = 10-3 мм) до 5 см и более, но в среднем от 5 до 250 мк. Самые мелкие среди них — внутриклеточные паразиты, например некоторые споровики и жгутиконосцы. Самые крупные представители — колониальные радиолярии, размеры которых могут достигать 25 см.
Подцарство Одноклеточные (Простейшие) объединяет животных, тело которых состоит из одной клетки. Она выполняет функции самостоятельного организма. Клетка простейшего состоит из цитоплазмы, органоидов, одного или нескольких ядер. В ней происходят обмен веществ с внешней средой, процессы размножения в развития.
Многие одноклеточные обладают специальными органоидами (движения, питания, выделения), возникшими как результат приспособления к среде обитания.
Клетка — это самовоспроизводящееся образование, отделённое от своего окружения плазматической мембраной, способствующей регуляции обмена между внутренней и внешней средой.
Среда обитания
Простейшие животные — процветающая и разнообразная группа (около 70 000 видов) — обитатели водоёмов и влажной почвы. Преимущественно они входят в состав зоопланктона — совокупность мельчайших животных, обитающих в морских и пресноводных водоёмах. На суше они встречаются тоже в водной среде — в почвенной капельной воде, а также в жидкой среде внутри многоклеточных животных и растений. Хотя почвенные простейшие животные могут существенно влиять на количество бактерий, все же их значение несравненно меньше, чем у простейших в пресных и морских водоёмах.
Жизнедеятельность
см. Питание простейших
Многие простейшие животные так же мелки и просто устроены, как и некоторые клетки крупных животных. Но они отличаются от них тем, что способны жить самостоятельно. Одноклеточные животные представляют собой слаженный организм, осуществляющий питание, дыхание, выделение, размножение, рост, развитие и обмен веществ. У него в протоплазме существует как бы разделение труда: каждая из её обособленных, более мелких образований выполняет свою особую задачу.
Например, ядро регулирует жизнедеятельность всего одноклеточного организма и воспроизводит само себя, благодаря чему образуются новые дочерние организмы; в пищеварительной вакуоли происходит переваривание пищи; сократительная вакуоль удаляет избыток воды и растворенные в ней вредные для организма вещества.
При неблагоприятных условиях многие простейшие перестают питаться, теряют органы движения, покрываются толстой оболочкой и образуют цисту. При наступлении благоприятных условий одноклеточные принимают прежний облик.
Классификация
Согласно названию Protozoa, в это подцарство должны входить только животные. Но в современной системе простейших содержатся зелёные жгутиконосцы (ботаники считают их водорослями), миксомицеты и плазмодиофориды (по мнению микологов, это грибы) и т. д. В связи с этим древние простейшие скорее всего могут рассматриваться в качестве исходной группы, давшей начало и грибам, и растениям, и животным. Поэтому в настоящее время должно считаться признанным выделение особого царства протистов и противопоставление его царствам растений и животных. Выделение царства протистов принадлежит знаменитому зоологу и эволюционисту Э. Геккелю (1866). Protozoa же могут быть выделены в системе протистов в качестве подцарства.
Одноклеточные прошли длительный путь эволюции, в ходе которой возникло их огромное разнообразие. В зависимости от сложности строения и способов передвижения выделяют несколько типов простейших. Материал с сайта http://wikiwhat.ru
Значение и роль
Со времён Линнея и до наших дней простейшие привлекают внимание учёных по разным причинам. Возникла даже специальная наука — протозоология.
Модель клетки
Простейшие представляют особый уровень развития живого мира поэтому их изучение имеет общебиологическое значение. В настоящее время простейшие используются как модель эукариотной клетки для исследований в области молекулярной биологии, генетики, радиобиологии и других наук.
Роль в биосфере
Простейшие играют важную роль в биосфере, являясь необходимым звеном в круговороте веществ и потоке энергии. Простейшие животные — очень важные потребители бактерий и одноклеточных водорослей, а также животных, в основном одноклеточных. По объёму потребляемого органического вещества они занимают третье место вслед за бактериями и грибами. Они образуют начальные звенья в сетях питания после растительных организмов — первичных продуцентов органических веществ.
Возбудители заболеваний
Одноклеточные являются возбудителями заболеваний человека и домашних животных.
Вопросы к этой статье:
Расскажите о разнообразии простейших.
Кто из представителей простейших имеет наиболее сложное строение?
Каково значение простейших в природе и жизни человека?
В чём заключается средообразующая роль простейших?
Простейшие [Одноклеточные] — животное, описание, характеристика, строение, питание, дыхание, размножение, где обитает, виды, фото, вики — WikiWhat
История открытия
Как известно, в 1675 г. т. е. более трёхсот лет назад, А. Левенгук открыл «анималькулов» (зверушек), которых впоследствии назвали инфузориями. С 1820 г. установилось название Protozoa, что в переводе с греческого означает «простейшие животные». Зоолог К. Зибольд посчитал их особым типом животного царства и выделил два класса: инфузорий и корненожек. Он же определил, что простота их организации соответствует одной клетке. С тех пор одноклеточность простейших стала общепризнанной, а название «одноклеточные» и «простейшие» стали синонимами.
По уровню организации все живые организмы классифицируются на две группы. Привычное для нас деление на одноклеточных и многоклеточных потребовало уточнения, после того как при изучении строения организмов был применён электронный микроскоп и появились новые методы исследования. Возникли вопросы об основных различиях, определяющих уровни развития, а также о планах строения. Поэтому необходимо рассмотреть организацию простейших — парафилетической группы, объединяющей представителей органического мира, относимых ранее к растениям, животным и грибам, но имеющих свои специфические особенности.
Происхождение
Самозарождение
Природа простейших долгое время оставалась предметом спора. Одни учёные рассматривали их как живых молекул, или простые комплексы таких молекул, которые способны самозарождаться, т. е. возникать сами по себе. Этих воззрений придерживались немногие учение, тем более что блестящие опыты Л. Спаланцани в XVIII в. Л. Пастера в XIX в. опровергли идею самозарождения.
Целлюляризация
Другие учёные считали простейших весьма сложно организованными существами, которых можно структурно сравнить с высокоорганизованными животными. Основание для этого они видели в том, что в организме многоклеточных есть структуры, не имеющие разделения на клетки, например синцитии. Исходя из подобных воззрений, зоолог Й. Хаджи в 50-60-е годы XX в. выдвинул даже теорию происхождения многоклеточных животных путём целлюляризации. Обнаружив сходство инфузорий с самыми примитивными ресничными червями, так называемыми бескишечными, Хаджи предположил, что при обособлении частей тела инфузории, содержащих органоиды, и образовании между ними перегородок возникает многоклеточный организм. Следовательно, по своей природе инфузория сравнима с целым организмом низших многоклеточных. Однако после электронно-микроскопических исследований было доказано, что теория целлюляризации опирается только на внешние аналогии и конвергентные сходства.
Клеточная теория Т. Шванна
С позиций клеточной теории, разработанной М. Шлейденом и Т. Шванном, простейшие представляют собой одноклеточные организмы. По мнению современных учёных, придерживающихся этих воззрений, простейшие — это клетки, которые функционально являются организмами. Однако функции не могут существовать отдельно от определённых структур. Таким образом, современное определение простейших как микроскопических одноклеточных животных, представляющих собой физиологически самостоятельные организмы, не соответствует нынешнему уровню знаний. Удовлетворительное определение простейших может быть дано после ответов на следующие вопросы: являются ли простейшие только одноклеточными организмами? Всегда ли их размеры микроскопически малы? Являются ли они исключительно животными? Являются ли они организмами только в физиологическом отношении?
Размеры простейших варьируют в той же степени, что и размеры многоклеточных. В среднем они колеблются от 5 до 250 мкм. Наиболее мелки внутриклеточные паразиты, например лейшмании (1-4 мкм). Но уже инфузория-туфелька значительно крупнее — 150-250 мкм, амёба протей — 600 мкм, а современные глубоководные радиолярии имеют диаметр тела 2,5 см. В современную систему простейших входят миксомицеты — настоящие гиганты среди протистов. Длина плазмодия одного из них доходит до 1,2 м. Короче говоря, различия в размерах простейших равняются 106.
Раньше считалось, что все простейшие — микроскопические существа. Сегодня среди них обнаружены и макроскопические виды. Размеры простейших колеблются от 1 мк (1 мк = 10-3 мм) до 5 см и более, но в среднем от 5 до 250 мк. Самые мелкие среди них — внутриклеточные паразиты, например некоторые споровики и жгутиконосцы. Самые крупные представители — колониальные радиолярии, размеры которых могут достигать 25 см.
Подцарство Одноклеточные (Простейшие) объединяет животных, тело которых состоит из одной клетки. Она выполняет функции самостоятельного организма. Клетка простейшего состоит из цитоплазмы, органоидов, одного или нескольких ядер. В ней происходят обмен веществ с внешней средой, процессы размножения в развития.
Многие одноклеточные обладают специальными органоидами (движения, питания, выделения), возникшими как результат приспособления к среде обитания.
Клетка — это самовоспроизводящееся образование, отделённое от своего окружения плазматической мембраной, способствующей регуляции обмена между внутренней и внешней средой.
Среда обитания
Простейшие животные — процветающая и разнообразная группа (около 70 000 видов) — обитатели водоёмов и влажной почвы. Преимущественно они входят в состав зоопланктона — совокупность мельчайших животных, обитающих в морских и пресноводных водоёмах. На суше они встречаются тоже в водной среде — в почвенной капельной воде, а также в жидкой среде внутри многоклеточных животных и растений. Хотя почвенные простейшие животные могут существенно влиять на количество бактерий, все же их значение несравненно меньше, чем у простейших в пресных и морских водоёмах.
Жизнедеятельность
см. Питание простейших
Многие простейшие животные так же мелки и просто устроены, как и некоторые клетки крупных животных. Но они отличаются от них тем, что способны жить самостоятельно. Одноклеточные животные представляют собой слаженный организм, осуществляющий питание, дыхание, выделение, размножение, рост, развитие и обмен веществ. У него в протоплазме существует как бы разделение труда: каждая из её обособленных, более мелких образований выполняет свою особую задачу.
Например, ядро регулирует жизнедеятельность всего одноклеточного организма и воспроизводит само себя, благодаря чему образуются новые дочерние организмы; в пищеварительной вакуоли происходит переваривание пищи; сократительная вакуоль удаляет избыток воды и растворенные в ней вредные для организма вещества.
При неблагоприятных условиях многие простейшие перестают питаться, теряют органы движения, покрываются толстой оболочкой и образуют цисту. При наступлении благоприятных условий одноклеточные принимают прежний облик.
Классификация
Согласно названию Protozoa, в это подцарство должны входить только животные. Но в современной системе простейших содержатся зелёные жгутиконосцы (ботаники считают их водорослями), миксомицеты и плазмодиофориды (по мнению микологов, это грибы) и т. д. В связи с этим древние простейшие скорее всего могут рассматриваться в качестве исходной группы, давшей начало и грибам, и растениям, и животным. Поэтому в настоящее время должно считаться признанным выделение особого царства протистов и противопоставление его царствам растений и животных. Выделение царства протистов принадлежит знаменитому зоологу и эволюционисту Э. Геккелю (1866). Protozoa же могут быть выделены в системе протистов в качестве подцарства.
Одноклеточные прошли длительный путь эволюции, в ходе которой возникло их огромное разнообразие. В зависимости от сложности строения и способов передвижения выделяют несколько типов простейших. Материал с сайта http://wikiwhat.ru
Значение и роль
Со времён Линнея и до наших дней простейшие привлекают внимание учёных по разным причинам. Возникла даже специальная наука — протозоология.
Модель клетки
Простейшие представляют особый уровень развития живого мира поэтому их изучение имеет общебиологическое значение. В настоящее время простейшие используются как модель эукариотной клетки для исследований в области молекулярной биологии, генетики, радиобиологии и других наук.
Роль в биосфере
Простейшие играют важную роль в биосфере, являясь необходимым звеном в круговороте веществ и потоке энергии. Простейшие животные — очень важные потребители бактерий и одноклеточных водорослей, а также животных, в основном одноклеточных. По объёму потребляемого органического вещества они занимают третье место вслед за бактериями и грибами. Они образуют начальные звенья в сетях питания после растительных организмов — первичных продуцентов органических веществ.
Возбудители заболеваний
Одноклеточные являются возбудителями заболеваний человека и домашних животных.
Вопросы к этой статье:
Расскажите о разнообразии простейших.
Кто из представителей простейших имеет наиболее сложное строение?
Каково значение простейших в природе и жизни человека?
В чём заключается средообразующая роль простейших?
Значение простейших в природе и жизни человека
Простейшие — источник питания для других животных. В морях и в пресных
водах простейшие, прежде всего инфузории и жгутиковые, служат пищей для
мелких многоклеточных животных. Черви, моллюски, мелкие ракообразные, а
также мальки многих рыб питаются преимущественно одноклеточными. Этими
мелкими многоклеточными, в свою очередь, питаются другие, более крупные
организмы. Самое большое из когда-либо живших на Земле животных — голубой
кит, как и все другие усатые киты, питается очень мелкими ракообразными,
населяющими океаны. А эти рачки питаются одноклеточными организмами. В
конечном счете существование китов зависит от одноклеточных животных и
растений.
Простейшие — участники образования горных пород. Рассматривая под микроскопом
размельченный кусочек обыкновенного писчего мела, можно видеть, что он
состоит преимущественно из мельчайших раковинок каких-то животных. Морские
простейшие (корненожки и радиолярии) играют весьма важную роль в образовании
морских осадочных горных пород. В течение многих десятков миллионов лет
их микроскопически мелкие минеральные скелеты оседали на дно и образовывали
мощные отложения. В древние геологические эпохи при горообразовательном
процессе морское дно становилось сушей. Известняки, мел и некоторые другие
горные породы в значительной мере состоят из остатков скелетов морских
простейших. Известняки с давних пор имеют огромное практическое значение
как строительный материал.
Изучение ископаемых остатков простейших играет большую роль в определении
возраста разных слоев земной коры и нахождении нефтеносных слоев.
Борьба с загрязненностью водоемов — важнейшая государственная задача.
Простейшие — показатель степени загрязненности пресных водоемов. Каждому
виду простейших животных необходимы для существования определенные условия.
Одни простейшие живут только в чистой воде, содержащей много растворенного
воздуха и не загрязненной отходами фабрик и заводов; другие приспособлены
к жизни в водоемах средней загрязненности. Наконец, есть и такие простейшие,
которые могут жить в очень загрязненных, сточных водах. Таким образом,
нахождение в водоеме определенного вида простейших дает возможность судить
о степени его загрязненности.
Простейшие — возбудители болезней человека и животных. Среди простейших
очень многие ведут паразитический образ жизни. Они поселяются в различных
органах человека и животных и часто бывают причиной тяжелых заболеваний.
К болезням, вызываемым простейшими, относятся, например, малярия и кожный
лейшманиоз (см. ст. «Насекомые и клещи — хранители и переносчики возбудителей
болезней»).
Итак, простейшие имеют огромное значение в природе и в жизни человека.
Одни из них не только полезны, но и необходимы; другие, напротив, опасны.
Простейшие-паразиты
☰
Среди простейших организмов есть множество паразитов животных и человека. Ниже перечислены паразитические простейшие, вызывающие наиболее серьезные и часто встречающие заболевания.
Дизентерийная амеба
Дизентерийная амеба похожа на обыкновенную, но мельче ее и имеет более короткие и широкие корненожки. В пищеварительную систему человека она попадает через рот в стадии цист. В толстом кишечнике амеба выходит из цисты и питается бактериями, не нанося вреда человеку. В дальнейшем этот простейший организм начинает внедряться в стенку кишечника, питаться эритроцитами крови, и становится паразитом. В кишечнике образуются язвы, истощающие организм человека. Возникает заболевание амебная дизентерия, или амебиаз.
Дизентерийная амеба может попадать в кровяное русло и достигать печени. Здесь также паразит приводит к образованию гнойных язв.
Образуя цисты, амебы покидают организм человека с непереваренными остатками пищи. Легкие цисты легко разносятся. Если не мыть руки и еду, то ими можно заразиться.
Малярийный плазмодий
Плазмодии представляют собой паразитических простейших. Некоторые виды плазмодиев вызывают у человека заболевание малярию. Переносчиком малярийных плазмодиев является малярийный комар. Во время укуса насекомого плазмодий проникает в кровь хозяина. Вместе с кровью он достигает печени, питается там, растет и размножается. После этого множество плазмодиев снова выходят в кровяное русло и начинают паразитировать на эритроцитах, разрушая их и выделяя свои продукты жизнедеятельности, которые отравляют хозяина. У человека начинается лихорадка, он страдает от малокровия.
Если больного малярией снова укусит малярийный комар, то теперь плазмодии попадут от человека к комару. В теле комара плазмодий размножается половым путем.
Малярия распространена в Африке. Это очень опасное заболевание. С малярией борются в том числе и уничтожая малярийных комаров.
Трипаносомы
Род трипаносомы составляют простейшие-паразиты, имеющие жгутики (по родству близкие к эвглене). Основной их хозяин — позвоночное животное, а переносчиками обычно являются насекомые. Разные представители трипаносом вызывают разные заболевания животных и человека. Паразитируют в основном в крови и спинномозговой жидкости. Самым известным и распространенным заболеванием, вызываемым видом трипаносом, является сонная болезнь.
Переносчиком сонной болезни является муха цеце. Данное заболевание характерно для тропической Африки. Сонная болезнь развивается в две стадии: первые недели человека мучает лихорадка и боли, через месяц или более наступает сонливость, нарушение сна и координации, изменение сознания. Болезнь легче лечится на первой стадии.
Лямблии
Лямблии — род жгутиковых простейших-паразитов. Кишечная лямблия вызывает у человека и животных лямблиоз, при котором паразит живет в тонкой кишке.
Человек заражается лямблиозом при поедании немытой пищи, содержащей цисты лямблии. Выйдя из цисты, лямблия присасывается к кишечнику и питается переваренной пищей.
Лейшмании
Лейшмании — это еще один род простейших-паразитов. Они вызывают лейшманиозы у человека и многих других животных. Переносчиками являются москиты.
Существуют различные виды лейшманиозов, связанные с поражением различных тканей организма. Одним из них является кожное заболевание пендинская язва.
Кокцидии
Кокцидии паразитируют на многих животных, в том числе червях, членистоногих, рыбах. Вызывают заболевания кокцидиозы, которые наносят серьезный урон в животноводстве и рыбоводстве.
Кокцидии расселяются в виде спор, содержащих клетки паразитов.
К кокцидиям относится род Токсоплазмы. Их представители вызывают у человека такое широко распространенное заболевание как токсоплазмоз. Человек заражается от домашних питомцев или плохо приготовленной мясной пищи. Токсоплазмы поражают многие органы, в том числе нервную систему.
Разнообразие и экологическое значение простейших
Бак, Р.П.М. и Nieuwland, G. (1989) Сезонные колебания в популяциях донных простейших на разных глубинах в морских отложениях. Neth. J. Sea Res.
24 , 37–44.
Google Scholar
Бамфорт, С.С. (1985) Роль простейших в подстилках и почвах. J. Protozool.
32 , 404–9.
Google Scholar
Бернингер, У.-Г, Карон, Д.А., Сандерс, Р.В., Финлей, Б.Дж. (1991) Гетеротрофные жгутиконосцы планктонных сообществ, их характеристики и методы изучения. В Биология свободноживущих гетеротрофных жгутиконосцев (Д. Дж. Паттерсон и Дж. Ларсен, ред.), Стр. 39–56. (Специальный выпуск Ассоциации систематики № 45) Оксфорд: Clarendon Press.
Google Scholar
Бик, Х. (1972) Реснитчатые простейшие . Женева: Всемирная организация здравоохранения.
Google Scholar
Бак, К.Р., Чавес, Ф.П. и Томсен, Х.А. (1991) Хоанофлагелляты центральных вод Калифорнии: численность и распространение. Офелия
33 , 179–86.
Google Scholar
Canning, E.U. (1982) Оценка характеристик простейших в отношении биологической борьбы с вредителями. Паразитология
84 , 119–49.
Google Scholar
Карон, Д.А. и Goldman, J.C. (1990) Регенерация питательных веществ морскими простейшими. В Экология морских простейших (Г. М. Каприуло, редактор), стр. 283–306. Нью-Йорк: Издательство Оксфордского университета.
Google Scholar
Кливленд, Л.Р. (1924) Физиология и симбиотические отношения между кишечными простейшими термитов и их хозяином, со специальной ссылкой на Reticulitermes flavipes Kollar. Biol. Бык.
46 , 117–227.
Google Scholar
Коул, Дж. Дж., Финдли, С. и Пейс, М. Л. (1988) Бактериальное производство в пресноводных и морских экосистемах: обзор системы сельскохозяйственных культур. Mar. Ecol. Прог. Сер.
43 , 1–10.
Google Scholar
Корлисс, Дж. О. (1984) Королевство Протиста и его 45 типов. Биосистемы
17 , 87–126.
Google Scholar
Корлисс, Дж. О. и Даггетт, П.-М. (1983) «Paramecium aurelia» и «Tetrahymena pyriformis»: текущий статус таксономии и номенклатуры этих широко известных и широко используемых инфузорий. Protistologica
19 , 307–22.
Google Scholar
De Jonckheere, J.F. (1987) Характеристика видов Naegleria с помощью рестрикционного расщепления цельноклеточной ДНК эндонуклеазой. Мол. Биохим. Паразит.
24 , 55–66.
Google Scholar
Доллетт М. (1984) Болезни растений, вызываемые жгутиковыми простейшими. Ann. Преподобный путь растений.
22 , 115–32.
Google Scholar
Фенчел Т. (1986) Экология простейших . Берлин: Мэдисон / Спрингер Верлаг.
Google Scholar
Фенчел, Т.и Finlay, B.J. (1983) Частота дыхания у гетеротрофных свободноживущих простейших. Microb. Ecol.
9 , 99–122.
Google Scholar
Фенчел Т. и Финли Б. Дж. (1989) Кентрофорос : инфузория без рта с симбиотическим огородом. Офелия
30 , 75–93.
Google Scholar
Фенчел, Т. и Финли, Б.J. (1991) Биология свободноживущих анаэробных инфузорий. евро. J. Protistol.
26 , 210–5.
Google Scholar
Фойсснер В. (1987) Простейшие в почве: фундаментальные проблемы, экологическое значение, адаптации инфузорий и панцирей, биоиндикаторы и справочник по литературе. Прог. Протистол.
2 , 69–212.
Google Scholar
Гарфинкель, Л.J., Giladi, M., Humber, M., Gitler, C., Mirelman, D., Evel, M. и Rozenblatt, S. (1989) ДНК-зонды, специфичные для Entamoeba histolytica , обладающих патогенными и непатогенными зимодемами . Заражение. Иммун.
57 , 926–31.
Google Scholar
Грэм, Дж. М. (1991) Краткая история водной микробной экологии. J. Protozool.
38 , 66–9.
Google Scholar
Грант, Дж.и Швингхамер, П. (1987) Размерное распределение микробной и мейобентосной биомассы и дыхание на берегу Брауна, юго-запад Новой Шотландии. Estuar. Берег. Shelf Sci.
25 , 647–61.
Google Scholar
Kreier, J.P. (1977) Parasitic Protozoa . Нью-Йорк: Academic Press.
Google Scholar
Маргулис Л., Корлисс Дж.О., Мелконян М.и Чепмен, Д.Дж., ред. (1990) Справочник по Protoctista . Бостон: Джонс и Бартлетт.
Google Scholar
Мартин, М. (1991) Эволюция переваривания целлюлозы у насекомых. Фил. Пер. Royal Soc. Лондон, серия B
333 , 281–7.
Google Scholar
Мейерс, Т.Р. (1990) Болезни ракообразных: болезни, вызываемые простейшими.В Болезни морских животных (О. Кинне, ред.) Т. 3. С. 330–68. Гамбург: Biologische Anstalt Helgland.
Google Scholar
Паттерсон, Д.Дж. и Ларсен, Дж. (1991) (ред.) Биология свободноживущих гетеротрофных жгутиконосцев . (Специальный выпуск Ассоциации систематики № 45). Оксфорд: Clarendon Press.
Google Scholar
Филлипсон, Дж.(1983) Биоиндикаторы, биологический надзор и мониторинг. Verheit. Втор. Zool. Геселл.
198 , 121–3.
Google Scholar
Pomeroy, L.R. (1974) Пищевая сеть океана; меняющаяся парадигма. BioScience
24 , 499–504.
Google Scholar
Портер, К.Г., Шерр, Э.Б., Шерр, Б.Ф., Пейс, М.Л. и Сандарс, Р.(1985) Простейшие в планктонных пищевых сетях. J. Protozool.
32 , 409–15.
Google Scholar
Шлегель М. (1991) Эволюция протистов и филогения, выявленные при сравнении последовательностей малых субъединиц рибосомных РНК. евро. J. Protist.
27 , 207–19.
Google Scholar
Сметачек В. (1981) Годовой цикл протозоопланктона в Кильской бухте. Mar. Biol.
63 , 1–11.
Google Scholar
Стивенс, Дж. Р. и Годфри, Д. Г. (1992) Численная таксономия трипанозоидов основана на полиморфизмах сокращенного набора ферментов. Паразитология
104 , 75–86.
Google Scholar
Stoecker, D.K., Michaels, A.E. и Davis, L.H. (1987) Обнаружено, что большая часть морских планктонных инфузорий содержит функциональные хлоропласты. Природа
326 , 790–2.
Google Scholar
Tait, A. and Turner, C.M.R. (1990) Генетический обмен в Trypanosoma brucei . Паразит. Сегодня
6 , 70–5.
Google Scholar
Тибайренк М. и Аяла Ф.Дж. (1991) К популяционной генетике микроорганизмов: клональная теория паразитических простейших. Паразит.Сегодня
7 , 228–32.
Google Scholar
Викерман, К. (1986) Тайный секс в трипаносомах. Природа
322 , 113–4.
Google Scholar
Виноградова Н.Г., Левенштейн Р.Ю. и Турпаева, Е. (1978) Количественное распределение донной фауны в районе исследований 16-го рейса научно-исследовательского судна «Дмитрий Менделеев». Труды Института Океанологии
113 , 7–21.
Google Scholar
Уильямс А.Г. (1991) Биохимическая активность и важность инфузорий простейших в экосистеме рубца. В Biochemical Protozoology (Дж. Х. Кумбс и М. Дж. Норт, ред.), Стр. 61–79. Лондон: Тейлор и Фрэнсис.
Google Scholar
Простейшие паразиты — Фонд безопасной питьевой воды
Что такое простейшие паразиты?
Простейшие паразиты могут показаться сложными, но на самом деле их нетрудно понять.Чтобы упростить изучение этих одноклеточных организмов, мы рассмотрим каждую часть по отдельности — сначала простейшие, затем паразиты, а затем объединим их. Прочитав всю эту информацию, вы обязательно станете экспертом! Давайте начнем!
Что такое простейшие?
Простейшие — это то, что мы называем эукариотическим организмом, потому что это клетка, которая содержит истинное ядро и ограничена ядерной мембраной. Он состоит всего из одной клетки и настолько мал, что мы обычно не можем увидеть его без микроскопа.Группа простейших очень разнообразна и насчитывает около 50 000 членов, каждый со своими личными характеристиками — почти так же, как каждый человек отличается от другого.
Простейшие можно найти почти в каждом месте на Земле, которое вы только можете себе представить. От самой глубокой части океана до вершины самой высокой горы встречаются простейшие. Их можно найти в прудах, ручьях, реках, болотах и в большинстве почв. Даже самая загрязненная вода содержит некоторые виды простейших.
У простейших есть жизненный процесс, подобный животному, потому что они могут добывать себе пищу, поглощая частицы пищи из окружающей среды, а не производя свою собственную пищу путем фотосинтеза.Фотосинтез используется растениями для поглощения энергии солнечного света и преобразования поглощенной энергии в органические соединения, которые потребляются в пищу. Напротив, простейшие обычно получают питание, поглощая (проглатывая и / или заключая) частицы пищи и небольшие частицы питательных веществ, таких как бактерии, водоросли или даже другие простейшие.
Большинство простейших, живущих в окружающей среде, не вредны, за исключением простейших, вызывающих болезни, о которых мы скоро поговорим. Многие виды простейших даже полезны для окружающей среды, потому что помогают сделать ее более продуктивной.Они улучшают качество воды, поедая бактерии и другие частицы.
Что такое паразит?
Паразит — это организм, который получает пищу и укрытие на или в другом организме, но ничего не дает для выживания хозяина. Например, паразит в нашем теле будет жить за счет наших клеток, нашей энергии или пищи, которую мы едим, и, как следствие, лишать нас питания и энергии, которые нам необходимы для того, чтобы быть активными и оставаться здоровыми. Когда в нашем теле есть паразиты, мы подвержены риску болезней, болезней и различных проблем со здоровьем.
Разве моя иммунная система не избавится от паразита, если он в моем теле? Хотя ответ на этот вопрос положительный, это не так просто, как распознать паразита, а затем устранить его. Паразиты — штука хитрая. Они могут жить в наших телах, а мы даже не подозреваем, что они там есть! Умный паразит живет незамеченным, чтобы оставаться там сколь угодно долго. Они способны воспроизводиться и выживать прямо под носом иммунной системы, оставаясь незамеченными. Как они это делают, остается загадкой…
Паразиты, как и простейшие, живут почти везде и могут легко проникнуть в нас разными способами.Вот некоторые из этих способов: укусы насекомых, хождение без обуви или носков, употребление грязной воды, употребление сырых фруктов и овощей и прикосновение к чему-то грязному, например, дверной ручке или деньгам, а затем прикосновение к нашему лицу или рту. Когда дело доходит до предотвращения паразитов, цель игры — осторожность.
Итак, мы узнали о простейших и узнали о паразитах. Давайте соберем их вместе и выясним, что такое простейшие паразиты.
Что такое простейший паразит?
Простейшие паразиты — это, в основном, простейшие, которые приспособились проникать в клетки и ткани других организмов и жить в них.Можно сказать, что простейшие усвоили уроки у паразитов, чтобы узнать, как они живут и выживают, а затем медленно начали меняться, чтобы стать более похожими на них. Простейшие превратились из способности выжить в одиночестве в окружающей среде, чтобы ухаживать за другими. Простейшие паразиты не могут жить в открытой среде сами по себе, как другие простейшие, но должны вторгаться в организм другого организма, чтобы получить защиту и питание.
Есть несколько простейших паразитов, вызывающих проблемы. Они присутствуют в нашей пище, почве и воде и могут вызвать у нас серьезное заболевание, если когда-нибудь проникнут внутрь нас.Каждый отдельный простейший паразит может вызывать разные болезни или болезни в нашем организме. Некоторые из них могут вызывать серьезные заболевания дыхательных путей (дыхательные пути из носа в легкие) и центральной нервной системы (головного мозга, черепных нервов и спинного мозга), в то время как другие живут в нашем кишечнике и вызывают такие симптомы, как диарея, и не являются смертельными. Некоторыми примерами несмертельных простейших патогенов являются Cryptosporidium, Giardia и Entamoeba histolytica.
Биология почвы: ключевые образовательные сообщения
Используйте этот план для организации биологии почвы.
презентация или определение ключевых моментов для публикации в печатных материалах.
Почва живая!
- В большинстве экосистем больше жизни и
разнообразие живет под землей, чем наверху.- Энергия течет от солнца через
растений, а также через многие трофические уровни почвенных организмов.- Почвенные организмы можно разделить на шесть
группы: бактерии, грибы, простейшие, нематоды, членистоногие и дождевые черви. Каждая группа
организмов играет важную роль.Даже внутри каждой группы существует большое разнообразие форм.
и функция.- Ризосфера — это интерфейс между
корни растений и почвенная среда. Это место большой биологической активности почвы.
и взаимодействия растений с микробами, включая симбиозы, патогенные инфекции и конкуренцию.- Почвенные организмы являются частью живого
система. Характеристики экосистемы во многом определяют структуру почвы.
сообщества.Погода определяет суточные и сезонные колебания биологической активности.
- Типы существующих видов и их
уровень активности зависит от микроклиматических условий, включая температуру,
влажность, аэрация, pH, размер пор и типы источников пищи.- В засушливых системах дождевых червей мало, но
есть термиты, муравьи и другие беспозвоночные, которые выполняют аналогичные функции.- Пастбища имеют примерно равное количество
грибковая и бактериальная биомасса, или в ней могут преобладать бактерии.Хвойные леса могут иметь
В 100-1000 раз больше биомассы грибов, чем биомассы бактерий.
Нам нужны почвенные организмы для оказываемых ими услуг. Они играют решающие роли
в здоровье растений и водной динамике.
- Почвенные организмы являются неотъемлемой частью
круговорот питательных веществ в окружающей среде. Едут:
- Разложение,
- Минерализация (E.g., простейшие и
нематоды выделяют избыток азота в почву, когда они поедают богатые азотом бактерии и
грибы.)- Хранение и выделение питательных веществ,
- Разложение загрязняющих веществ до того, как они
достигли грунтовых или поверхностных вод,- Круговорот углерода, секвестрация углерода и
трансформации органического вещества почвы,- Круговорот азота (фиксация азота,
денитрификация, нитрификация).
- Существенная биологическая активность почвы
влияет на структуру почвы, включая размер почвенных пор, стабильность почвенных агрегатов,
и наличие макропор. Структура почвы влияет на то, как вода перетекает, внутрь и
через почву и сколько воды удерживается в пределах досягаемости корней растений.
- Крупные роющие беспозвоночные (например,
дождевые черви, муравьи, термиты, жуки) образуют макропоры, которые обеспечивают быстрое попадание воды в
или через почву.- Даже крошечные членистоногие производят фекалии
пеллеты, представляющие собой смесь почвы и органических веществ. Они стали устойчивыми агрегатами почвы.- Грибы и бактерии производят вещества
которые помогают связывать частицы почвы и стабилизировать агрегаты почвы.- Органическое вещество почвы физически может быть
защищен от деградации в стабильных почвенных агрегатах.
- Динамика численности вредителей растений зависит от целого
смесь организмов в почве.Некоторые организмы охотятся или конкурируют с болезнетворными
организмы. Некоторые бактерии выделяют факторы роста растений, которые непосредственно ускоряют рост растений.- На засушливых землях биологические корки почвы
кажутся важными для всех перечисленных выше целей. Они помогают фиксировать азот, стабилизировать
поверхность почвы, влияют на поток воды и предотвращают появление каких-то экзотических растений
разновидность.- Микоризные грибы помогают растениям приобретать
питательные вещества из почвы, и они помогают стабилизировать почвенные агрегаты.- Устойчивость — это способность почвы к
восстанавливают свои функции после таких помех, как пожар, уплотнение, обработка почвы и т. д.
организмов в почве частично определяют ее сопротивляемость.
Управление воздействует на почвенные организмы
- Мы знаем, что практика землепользования
изменить почвенное сообщество. Связь между конкретными изменениями и функцией почвы меньше
Чисто.- Биологические корки почвы очень чувствительны
топтать.- Уменьшение обработки почвы обычно приводит к
усиление роста грибков, в том числе микоризных грибов.- Уплотнение почвы, отсутствие растительности или
отсутствие подстилки, покрывающей поверхность почвы, приводит к уменьшению количества почвы
членистоногие.
Краткие сведения о биологии почвы
Кончики маленьких корней растений проходят сквозь
почву закручивающим винтообразным движением.На зрелых деревьях может быть до 5 штук.
миллион активных корневых кончиков.
Одна лопата, полная плодородной садовой почвы
содержит больше видов организмов, чем можно найти над землей на всей территории Амазонки.
тропический лес.
Хотя поверхность почвы кажется твердой,
воздух свободно входит и выходит из него. Воздух в верхних 8 дюймах хорошо дренированной почвы
полностью обновляется примерно каждый час.
Растения на пшеничном поле площадью 2 акра
может иметь более 30 000 миль корней, больше, чем окружность Земли.
Чудесный «землистый» запах
недавно вспаханная земля, как полагают, является результатом химикатов, производимых
микроорганизмы. Одно из этих химикатов, называемое геосмин, производится
актиномицеты, организмы, которые обладают некоторыми свойствами как бактерий, так и грибов.
Почва может действовать как сток или источник
парниковых газов. Примерно 30 процентов углекислого газа, 70 процентов
метан и 90 процентов закиси азота, выбрасываемой в атмосферу каждый год, проходят
через почву.
Требуется от 4000 до 6000 фунтов
пожнивные остатки в год для поддержания содержания органических веществ в почве.
Современные методы ведения сельского хозяйства, минимизирующие почву
нарушение (вспашка) и возвращение растительных остатков в почву, например, при нулевой обработке почвы и
севообороты медленно восстанавливают запасы органических веществ в почве.
углерода, возвращенного в почву в виде растений
остатков, от 5 до 15 процентов связывается в телах организмов, а от 60 до 75 процентов.
процентов вдыхается в виде углекислого газа обратно в атмосферу.Только от 10 до 25 процентов
превращается в перегной в почве.
«Мы знаем больше о движении
небесные тела, чем земля под ногами ».
— Леонардо да Винчи, около 1500-х гг.
«Каждый раз, когда вы делаете шаг в
зрелый лес Орегона, вашу ногу поддерживают 16000 беспозвоночных
удерживается в среднем 120 000 ног ».
— Д-р. Эндрю Молденке, Государственный университет Орегона.
Даже на сельскохозяйственных почвах более
тысячи ног членистоногих поддерживают каждый ваш шаг.
Одна чашка почвы может вместить столько бактерий
как есть люди на Земле.
Бактерии и актиномицеты чрезвычайно
крошечный. Тем не менее, из-за их огромной численности они составляют половину живой биомассы в
некоторые почвы.
Вес всех бактерий на одном акре
почвы может равняться весу коровы или двух.
Актиномицеты имеют клетки, подобные бактериям, но
растут такими же длинными нитями, как грибы. Как и грибы, они способствуют разложению твердых материалов, но
в отличие от грибов предпочитают высокий pH (более 7.0).
Чайная ложка сельскохозяйственной почвы может содержать десятки
ярды грибов. Такое же количество почвы из хвойного леса может вместить десятки миль
грибы.
Нематоды удивительно разнообразны. Двадцать
были описаны тысячи видов, но считается, что может существовать 500 000 видов.
Описано пять тысяч видов почв.
Дождевые черви перемещают почву из нижних слоев вверх
на поверхность и перемещают органические вещества с поверхности почвы в нижние слои. Где
дождевые черви активны, они могут перевернуть верхние 6 дюймов почвы за десять-двадцать лет.
<Вернуться в главное меню>
простейших | Что такое микробиология?
Простейшие — одноклеточные организмы. Они бывают разных форм и размеров, от Amoeba , который может менять свою форму, до Paramecium с фиксированной формой и сложной структурой. Они живут во множестве влажных мест обитания, включая пресную воду, морскую среду и почву.
Некоторые из них являются паразитическими, что означает, что они обитают в других растениях и животных, включая людей, и вызывают болезни.Например, Plasmodium, вызывает малярию. Они подвижны и могут перемещаться по:
- Реснички — крошечные, похожие на волоски структуры, покрывающие микроб снаружи. Они бьют равномерно, непрерывно, как гибкие весла.
- Жгутики — длинные нитевидные образования, отходящие от поверхности клетки. Жгутики движутся подобно хлысту, создавая волны, толкающие микроб.
- Амебоидное движение — организм движется, посылая псевдоподии, временные выступы, которые заполняются цитоплазмой, вытекающей из тела клетки.
© Фрэнк Фокс / Библиотека научных фотографий
Стентор — это простейшее в форме трубы, с кольцом из ресничек вокруг устья трубной «воронки». Реснички переносят бактерии и добычу, как мелких ракообразных, в это отверстие и вниз в пищеварительную систему.
О мышах и людях: миллиарды людей стали жертвами контроля над разумом токсоплазмы?
Toxoplasma gondii безмолвно пребывает в головах миллиардов людей по всему миру.В течение долгого времени заражение T. gondii считалось безвредным, но недавние данные свидетельствуют о том, что он модулирует уровни нейротрансмиттеров, изменяя личность и поведение, чтобы увеличить возможность его дальнейшей передачи.
Брифинг по политике в отношении малярии
Малярия — это инфекционное заболевание, вызываемое одноклеточным паразитарным организмом Plasmodium , который поражает кровь и печень.
Друзья с выгодой или эксплуатацией?
Эндосимбиозы, при которых один вид обитает внутри другого, встречаются повсюду в микробиологии. Например, зооксантеллы — это простейшие, обитающие внутри кораллов, морские беспозвоночные, которые строят коралловые рифы.
Древний остаток внутри малярийного паразита
Малярия вызывается простейшими паразитами из рода Plasmodium .Эти паразиты передаются через укусы комаров, и известно, что несколько различных видов заражают людей. Но загляните внутрь самой клетки Plasmodium , и вы обнаружите кое-что довольно неожиданное — клеточную структуру, которая очень похожа на хлоропласт.
Простейшие: патогенез и защита — Медицинская микробиология
Общие концепции
Устойчивость
Устойчивость — это способность хозяина защищаться от патогена.Устойчивость к простейшим паразитам включает три взаимосвязанных механизма:
неспецифические факторы, клеточный иммунитет и гуморальный иммунитет.
Патология
Протозойная инфекция приводит к повреждению тканей, ведущему к заболеванию. В хроническом
инфекции повреждение тканей часто происходит из-за иммунного ответа на паразита
и / или к антигенам хозяина, а также к изменениям профилей цитокинов.
В качестве альтернативы, это может быть связано с токсичными протозойными продуктами и / или механическим воздействием.
повреждать.
Механизмы побега
Механизмы побега — это стратегии, с помощью которых паразиты избегают убивающего эффекта
иммунная система иммунокомпетентного хозяина.Механизмы эвакуации, используемые
к простейшим паразитам относятся следующие.
Антигенная маскировка: Антигенная маскировка — это способность паразита к
избежать иммунного обнаружения, покрываясь антигенами хозяина.
Блокировка факторов сыворотки: Некоторые паразиты приобретают покрытие из
комплексы антиген-антитело или нецитотоксические антитела, которые стерически блокируют
связывание специфических антител или лимфоцитов с поверхностью паразита
антигены.
Внутриклеточное расположение: Внутриклеточная среда обитания некоторых простейших.
паразиты защищают их от прямого воздействия иммунного ответа хозяина.Скрывая антигены паразита, эта стратегия также задерживает обнаружение
иммунная система.
Антигенная вариация: Некоторые простейшие паразиты изменяют свою поверхность
антигены во время инфекции. Паразиты, несущие новые антигены
избежать иммунного ответа на исходные антигены.
Иммуносупрессия: Паразитарные простейшие инфекции обычно вызывают
некоторая степень иммуносупрессии хозяина. Этот сниженный иммунный ответ может задержать
обнаружение антигенных вариантов.Это также может снизить способность иммунной системы.
система для подавления роста и / или уничтожения паразитов.
Введение
Устойчивость к паразитическим простейшим похожа на устойчивость к другим
инфекционных агентов, хотя механизмы устойчивости к простейшим инфекциям
еще не так хорошо понят. Сопротивление можно разделить на две основные группы:
механизмы: (1) неспецифический механизм (ы) или фактор (ы), такие как наличие
неспецифический компонент сыворотки, смертельный для паразита; и (2) специфические
механизм (ы) с участием иммунной системы ().Вероятно, наиболее изученные неспецифические механизмы, участвующие в
резистентность к паразитам — это те, которые контролируют восприимчивость эритроцитов
к вторжению или росту плазмодиев, возбудителей малярии. Лица, которые
гетерозиготные или гомозиготные по серповидно-клеточному гемоглобину значительно
более устойчивы к Plasmodium falciparum , чем люди с
нормальный гемоглобин. Точно так же люди, у которых отсутствует фактор Даффи на красном
клетки крови не чувствительны к P vivax .Возможно, оба
серповидно-клеточный признак и отсутствие фактора Даффи были установлены в
эндемичные по малярии группы населения в результате избирательного давления, оказываемого малярией.
Эпидемиологические данные свидетельствуют о том, что другие наследственные аномалии эритроцитов,
такие как талассанемия и дефицит глюкозо-6-фосфатдегидрогеназы, может
способствуют выживанию людей в различных географических регионах, эндемичных по малярии.
регионы. Второй хорошо задокументированный пример неспецифического фактора, участвующего в
резистентность — это наличие в сыворотке крови человека трипанолитического фактора, который
придает устойчивость против Trypanosoma brucei brucei , возбудителя
трипаносомоз (сонная болезнь) у животных.Есть свидетельства того, что другие
неспецифические факторы, такие как лихорадка и пол хозяина, также могут способствовать
устойчивость хозяина к различным простейшим паразитам. Хотя неспецифические факторы
могут играть ключевую роль в сопротивлении, обычно они работают вместе с хозяином
иммунная система ().
Рисунок 78-1
Некоторые взаимосвязи между факторами хозяина, участвующими в устойчивости
к простейшим инфекциям.
Различные паразиты вызывают разные гуморальные и / или клеточные иммунные ответы.В
малярия и трипаносомные инфекции, антитела, по-видимому, играют важную роль в
иммунитет. Как в T cruzi , так и в T brucei gambiense
инфекции, антителозависимые цитотоксические реакции против паразита не наблюдались.
сообщил. Хотя было показано, что антитела ответственны за очистку
Последние данные свидетельствуют о том, что африканские трипаносомы из крови инфицированных животных
что время выживания инфицированных мышей не обязательно коррелирует с
способность животного вырабатывать антитела, специфичные для трипаносомы.Другими словами,
сопротивление, измеряемое временем выживания, может включать не только специфические гуморальные
иммунная система. Последние данные свидетельствуют о том, что для резистентности необходим клеточный иммунитет
малярии. например, испытания вакцины со спорозоитным антигеном показали, что
как активный клеточный ответ, так и антитела, специфичные для спорозоитов, могут потребоваться для
успешная иммунизация.
Считается, что клеточный иммунитет является самым важным защитным механизмом в
лейшманиоз и токсоплазмоз.У животных, инфицированных
Toxoplasma , было показано, что активированный макрофаг играет
важная роль в сопротивлении. Соответственно, устойчивость к простейшим паразитам
скорее всего, включает неспецифические факторы, а также специфические гуморальные и / или клеточные
механизмы. Цитокины участвуют в контроле как иммунного ответа, так и
патология. Стало очевидно, что существуют подмножества как помощника (h), так и
цитотоксические (c) Т-клетки, которые продуцируют разные профили цитокинов. Например,
Подмножество Th-1 продуцирует гамма-интерферон (IFN-α) и интерлейкин-2 (IL-2).
и участвует в клеточном иммунитете.Напротив, подмножество Th-2 продуцирует IL-4.
и IL-6, и отвечает за опосредованный антителами иммунитет. Индукция
конкретная подгруппа Т-клеток является ключом к выздоровлению и сопротивлению. Подмножество Th-1 и
повышенный уровень IFN-g важен для устойчивости к Leishmania ,
T cruzi и Toxoplasma , тогда как
Ответ Th-2 более важен при паразитарных инфекциях, в которых антитела являются ключевыми.
фактор. Важно понимать, что цитокины, продуцируемые одной Т-клеткой,
подмножество может усиливать или подавлять ответ других подмножеств Т-клеток.Ил-4 будет
подавляют активность клеток Th-1 и усугубляют инфекцию и / или восприимчивость мышей к
Лейшмания . Цитокины, продуцируемые Т и другими типами клеток,
не действуют непосредственно на паразитов, но влияют на другие типы клеток-хозяев. Ответ
клеток в цитокины включает в себя множество физиологических изменений, таких как изменения
в метаболизме глюкозы, жирных кислот и белков. Например, ИЛ-1 и некроз опухоли.
Фактор усилит глюконеогенез и окисление глюкозы. Следует отметить, что
цитокины влияют на метаболизм не только Т-клеток, но и множества других
типы клеток и системы органов.Цитокины также могут стимулировать деление клеток и,
следовательно, клональная экспансия подмножеств Т- и В-клеток. Это может привести к увеличению
продукция антител и / или количество цитотоксических Т-клеток. Список цитокинов и их
функций быстро растет, и кажется, что эти химические сообщения
влияют на все фазы иммунного ответа. они также явно участвуют в
наблюдается множество физиологических реакций (лихорадка, снижение потребления пищи и т. д.)
в реакции животного на патоген и в возникающей патологии.
В отличие от большинства вирусных и бактериальных инфекций, простейшие болезни часто бывают хроническими,
длящиеся месяцы или годы. Когда это связано с сильным иммунным ответом хозяина, это
Тип хронической инфекции может привести к высокому уровню иммунопатологии.
Также возникает вопрос, как эти паразиты выживают в иммунокомпетентном организме.
животное. В оставшейся части этой главы рассматриваются механизмы, ответственные за
патологии, в частности иммунопатологии, при простейших болезнях, и механизмы,
какие паразиты уклоняются от иммунных ответов хозяина.Наконец, из-за самого
быстрый прогресс в наших знаниях о взаимоотношениях паразит-хозяин (в первую очередь благодаря
развитие методик молекулярной биологии) необходимо кратко
упомянуть о возможности разработки вакцин против патогенных простейших.
Патология
Простейшие могут вызывать гуморальные ответы, при которых комплексы антиген-антитело в
область избытка антител активирует фактор свертывания крови Хагемана (фактор XII),
что, в свою очередь, активирует коагуляционную, фибринолитическую, кининовую и комплементарную системы.Было высказано предположение, что этот тип гиперчувствительности немедленного типа является причиной
при различных клинических синдромах африканского трипаносомоза, в том числе крови
повышенная вязкость, отеки и гипотония. Ожидаются аналогичные механизмы заболевания
при других инфекциях простейшими, вызывающими сильный гуморальный иммунный ответ ().
Таблица 78-1
Некоторые патологические механизмы протозойных заболеваний.
Иммунные комплексы циркулируют в сыворотке крови и откладываются в почках.
и другие ткани людей и животных, инфицированные простейшими.Эти паразиты
Комплексы антиген-антитело, плюс комплемент, были элюированы из почечной ткани в
случаи малярии и африканского трипаносомоза. Антиген и антитело были
непосредственно визуализируется в клубочках инфицированных животных с помощью света и электронов
микроскопия. Воспалительные клеточные инфильтраты сопровождают эти отложения и признаки
обычно наблюдаются гломерулонефриты. Африканские трипаносомы и предположительно их
антигены также обнаруживаются во множестве внесосудистых мест. Иммунные комплексы,
в этих тканях были обнаружены клеточные инфильтраты и повреждение тканей.
Другой важной формой патологии, опосредованной антителами, является аутоиммунитет. Аутоантитела
к ряду различных антигенов хозяина (например, эритроцитам, ламинину,
коллаген и ДНК). Эти аутоантитела могут играть роль в
патология паразитарных заболеваний двояко. Во-первых, антитела могут вызывать
прямое цитотоксическое действие на клетки-хозяева; например, аутоантитела, покрывающие красный
кровяные тельца вызывают гемолитическую анемию. Альтернативно аутоантитела могут быть
патогенны из-за накопления комплексов антиген-антитело в почках или других
тканей, что приводит к гломерулонефриту или другим формам гиперчувствительности немедленного типа.Особенно хороший пример простейшей инфекции, при которой проявляется аутоиммунитет.
вносить важный вклад в патогенез — T cruzi
инфекционное заболевание. В этом случае есть веские доказательства того, что хозяин и паразит имеют общие
перекрестно реагирующие антигены. Антитела и цитотоксические лимфоциты к этим антигенам
кажутся вредными для тканей хозяина. Этот тип экспериментальных данных в сочетании с
тот факт, что паразит как бы сам не вызывает патологии тканей, приводит
сделать вывод, что аутоиммунитет может играть ключевую роль в патогенезе.
Клеточная гиперчувствительность также наблюдается при простейших заболеваниях (). Например, при лейшманиозе
(вызвано Leishmania tropica ), поражения, по-видимому, вызваны
клеточно-опосредованный иммунный ответ и обладают многими, если не всеми, характеристиками
гранулемы, наблюдаемые при туберкулезе или шистосомозе. В этих поражениях
продолжающийся иммунный ответ на патогены, способные избежать защиты хозяина
механизмы вызывают дальнейший приток воспалительных клеток, что приводит к устойчивому
реакции и продолжающаяся патология в местах отложения антигена.Во время
паразитарная инфекция, различные продукты клетки-хозяина (цитокины, лимфокины и др.)
высвобождается из активированных клеток иммунной системы. Эти посредники влияют на
действие других клеток и может принимать непосредственное участие в патогенезе. Примером является
фактор некроза опухоли (TNF), который выделяется лимфоцитами. TNF может быть вовлечен
в мышечном истощении, наблюдаемом при хронических стадиях африканского трипаносомоза. TNF
также был причастен к кахексии и истощению в Leishmania
donovani инфекция, церебральная малярия в P falciparum
у детей и снижение выживаемости у мышей, инфицированных T cruzi- .это
очевидно, что медиаторы, участвующие в устойчивости к простейшим паразитам, также могут приводить к
патологии при хронической инфекции (). Похоже, существует тонкий баланс между факторами.
участвует в устойчивости к инфекционным агентам и тем, которые в конечном итоге вызывают
патология и клиника заболевания.
Многие авторы предположили, что токсичные продукты, продуцируемые паразитическими простейшими
несут ответственность хотя бы за некоторые аспекты патологии (). Например, гликопротеины на поверхности
было обнаружено, что трипаносомы фиксируют комплемент.Эта активация комплемента
предположительно приводит к производству биологически активного и токсичного комплемента
фрагменты. Кроме того, известно, что трипаносомы выделяют протеазы и
фосфолипазы, когда они лизируются. Эти ферменты могут вызывать разрушение клеток-хозяев,
воспалительные реакции и грубая патология тканей. Кроме того, это было
предположили, что трипаносомы содержат митоген В-клеток, который может изменять
иммунный ответ хозяина, вызывая поликлональный ответ B-клеток, который приводит к
иммуносупрессия.Наконец, недавно было показано, что африканские трипаносомы
также содержат эндотоксин, который предположительно высвобождается во время опосредованного антителами
лизис. Сообщалось также, что паразитические простейшие синтезируют (или содержат)
низкомолекулярные токсины. Например, трипаносомы производят несколько индол
катаболиты; в фармакологических дозах некоторые из этих катаболитов могут продуцировать
патологические эффекты, такие как лихорадка, вялость и даже подавление иммунитета. По аналогии,
ферменты, митоген В-клеток и т. д., по-видимому, высвобождаются многими, если не всеми,
другие паразитические простейшие.Работа по изучению роли этих простейших
продукты в патогенезе. Однако паразитические простейшие, как правило, не известны.
вырабатывают токсины с эффективностью, сопоставимой с классическими бактериальными токсинами
(например, токсины, вызывающие сибирскую язву и ботулизм). Одно возможное исключение:
африканские трипаносомы, которые предположительно содержат эндотоксин.
Immune Escape
Механизмы побега паразитов могут включать в себя ряд различных явлений (). При антигенной маскировке
паразит покрывается компонентами хозяина и поэтому не может быть распознан как
иностранный.При блокировании нецитотоксические антитела сочетаются с паразитарными антигенами и
подавляет связывание цитотоксических антител или клеток. Паразит может передавать часть
его жизненный цикл во внутриклеточном месте, например, в эритроцитах или
макрофаги, в которых он защищен от внутриклеточного пищеварения и от
цитотоксическое действие антител и / или лимфоцитов. Некоторые паразиты практикуют антигенные
вариации, изменяя свои поверхностные антигены в ходе инфекции и
таким образом избегая иммунных ответов хозяина.Наконец, паразит может вызвать
иммуносупрессия, снижение иммунного ответа хозяина на паразита
в частности или к чужеродным антигенам в целом. Эти стратегии обсуждаются в
подробнее ниже.
Таблица 78-2
Некоторые предлагаемые механизмы иммунного спасения, используемые простейшими
Паразиты.
Маскировка и мимикрия
Различные виды трипаносом имеют иммуноглобулины хозяина, связанные с их
поверхности ячеек. Есть несколько сообщений о том, что эти антитела не связываются с
трипаносомы через их вариабельные области, но предположительно через Fc
часть их молекулы.Эти антитела могут маскировать паразита, то есть
предотвратить иммунное распознавание хозяином. Однако никаких доказательств, кроме
наличие иммуноглобулинов на поверхности трипаносом подтверждает это
гипотеза. Мимикрия, при которой паразит имеет генетическую информацию для
синтезировать антигены, идентичные антигенам своего хозяина, не было продемонстрировано в
паразитические простейшие.
Блокирование
Было высказано предположение, что в некоторых случаях комплексы антиген-антитело в сыворотке
инфицированных животных связываются с поверхностью паразита, механически блокируя
действия цитотоксических антител или лимфоцитов и прямое ингибирование
действия лимфоцитов.Этот тип механизма иммунного ускользания был предложен
для опухолевых клеток и для паразитарных гельминтов. Потому что трипаносомы несут
иммуноглобулины на их клеточных поверхностях, они могут использовать аналогичный механизм;
однако никаких прямых доказательств пока не поступало.
Внутриклеточное расположение
Многие простейшие паразиты растут и делятся внутри клеток-хозяев. Например,
Plasmodium паразиты растут сначала в гепатоцитах, а затем в
красные кровяные клетки. Leishmania и Toxoplasma
организмы способны расти в макрофагах; один род паразитических
простейшие, Theilera , не только размножаются лимфоцитами, но и
кажется, даже стимулирует размножение инфицированных лимфоцитов.Хотя некоторые паразиты, такие как Plasmodium, ограничены в количестве
типов клеток-хозяев, другие, такие как T cruzi и
Toxoplasma , похоже, может расти и делиться в
множество различных клеток-хозяев.
Внутриклеточное убежище может защитить паразита от вредных или смертельных эффектов.
антител или клеточных защитных механизмов. Например,
Plasmodium может быть чувствительным к действию антител.
только во время коротких внеклеточных фаз своего жизненного цикла (спорозоит и
мерозоитовые стадии).Следует помнить, что Plasmodium
фактически находится в мембраносвязанной вакуоли в клетке-хозяине. Таким образом, плазмодии
защищены от внешней среды по крайней мере двумя мембранами хозяина (
внешняя мембрана клетки и внутренняя мембрана вакуоли). Хотя внутриклеточный
плазмодии очень хорошо защищены от иммунного ответа хозяина на ранних этапах своего развития.
рост, эта стратегия действительно создает физиологические проблемы для паразита. Для
Например, паразит должен получать питательные вещества для роста через три
мембраны (два хозяина и один паразит) и должны устранять продукты жизнедеятельности
через те же три мембраны.Плазмодии решают эту проблему соответствующим образом
модификация мембран клеток-хозяев. Паразитарные белки включены в состав
внешняя мембрана эритроцитов. Хозяин в конечном итоге реагирует на эти антигены,
и этот ответ в конечном итоге приводит к усиленному удалению инфицированного хозяина.
клетки.
Наличие внеклеточных фаз в жизненном цикле малярии важно,
поскольку иммунизация против этих стадий является основанием для развития
наши нынешние кандидаты на вакцины. Защитные антигены на этих внеклеточных
стадии были очищены как потенциальные антигены для вакцины.Однако это
подход имеет проблемы. Например, стадия спорозоитов подвергается воздействию
защитное антитело только на короткий период и даже один спорозоит, который
ускользает от иммунной элиминации, приведет к инфекции. Во-вторых, антигенный
изменчивость разных изолятов и способность разных штаммов к
подвергаются антигенной изменчивости, полностью не изучены. Следовательно, эффективность
вакцины-кандидаты все еще должны быть продемонстрированы. Однако большой синтетический
пептид, содержащий антигенные последовательности из 3 различных белков P
falciparum , снижает клиническую заболеваемость
малярия на 31% в полевых испытаниях.Поэтому есть оптимизм в отношении того, что вакцина
против P falciparum может быть доступен в ближайшем будущем.
Ряд паразитических простейших обитает в макрофагах. Хотя эти организмы
защищены от внешних иммунных угроз, они все равно должны избегать переваривания
макрофаг. Было предложено три стратегии. Во-первых, паразит может
предотвратить слияние лизосом с фагоцитарной вакуолью. Настоящий
механизм, ответственный за это торможение, еще не изучен, но он был
показано, что происходит в клетках, инфицированных Toxoplasma .Второй
Механизм представлен способностью T cruzi убегать
из фагоцитарной вакуоли в цитоплазму макрофага. Наконец, это
возможно, что некоторые паразиты могут выжить в присутствии лизосомальных ферментов,
как и лепрозная палочка. Один из наиболее изученных примеров простейших.
паразит, способный выжить в фаголизосоме — Leishmania . Это
Было высказано предположение, что устойчивость этого паразита к гидролитической
ферменты связаны с поверхностными компонентами, которые ингибируют ферменты хозяина и / или
наличие паразитарных ферментов, гидролизующих ферменты хозяина.В виде
ранее отмечалось, что по крайней мере один простейший паразит, Theilera , является
способен расти непосредственно в лимфоцитах. Следовательно, этот паразит может сбежать.
иммунный ответ хозяина за счет роста внутри самих клеток, необходимых для
отклик.
Антигенная вариация
Известно, что три основные группы паразитических простейших способны изменять
антигенные свойства их поверхностного покрытия. Африканские трипаносомы могут
полностью заменять антигены в их гликокаликсе каждый раз, когда хозяин проявляет
новый гуморальный ответ.Эти изменения серотипа являются одним из важных способов
африканские трипаносомы избегают защитного механизма хозяина. Хотя
менее хорошо охарактеризованные, аналогичные изменения происходят в
Плазмодий , Бабезия и
Лямблии.
Было подсчитано, что африканские трипаносомы имеют приблизительно 1000
разные гены, кодирующие поверхностные антигены. Эти гены расположены на разных
хромосомы; однако для экспрессии ген должен находиться в конце
хромосома (теломерный участок).Скорость, с которой происходит изменение
Популяция, передаваемая мухой цеце, довольно высока. Было показано, что 1
в 10 клетках, по-видимому, способен переключать свой поверхностный антиген. Порядок в
какие гены поверхностной оболочки экспрессируются, непредсказуемо. Много информации
имеется нуклеотидная последовательность генов, кодирующих белки оболочки;
однако ни фактор (ы), который побуждает клетку переключать свою поверхность
ни антигены, ни конкретный генетический механизм (ы), участвующие в переключении, полностью не
понял.Ответ антител не вызывает генетический переключатель, а просто
отбирает варианты с новыми поверхностными антигенами из исходной популяции.
О феномене антигенной активности известно значительно меньше.
вариации малярии или бабезиоза. Однако вариация антигена может быть основным
проблема, связанная с разработкой вакцины на стадии крови (мерозоит) для
малярия. Наконец, антигенная изменчивость наблюдалась у лямблий.
лямблия . Ряд различных семейств генов, кодирующих поверхностные
белки Giardia были идентифицированы.Антигенная вариация
было предложено помочь Giardia в побеге хозяина
иммунная реакция.
Иммуносупрессия
Иммуносупрессия хозяина наблюдалась почти у каждого паразита
Организм тщательно обследован на сегодняшний день. В некоторых случаях подавление носит специфический характер,
включающий только реакцию хозяина на паразита. В остальных случаях
подавление является гораздо более общим, включая реакцию на различные гетерологичные
и непаразитарные антигены. Еще не доказано, что эта иммуносупрессия
позволяет паразитам выжить в нормально иммунокомпетентном хозяине.Однако один
можно предположить, что иммуносупрессия может позволить небольшому количеству паразитов размножаться.
избежать иммунного надзора, способствуя тем самым возникновению хронической инфекции.
Этот механизм может быть особенно эффективным у паразитов, подверженных заражению.
антигенная вариация, так как это может позволить небольшому количеству паразитов с новыми
поверхностные антигены поначалу остаются необнаруженными. Иммуносупрессия экспериментально
индуцированные различными посторонними агентами, безусловно, вызывают более высокие
паразитемии, более высокий уровень инфицирования или и то, и другое.Следовательно, гипотеза о том, что
Иммосупрессия, вызванная паразитами, увеличивает вероятность заражения паразитом.
завершить его жизненный цикл имеет смысл.
Следует отметить, что иммуносупрессия сама по себе может быть патогенной. Сниженный
ответ на гетерологичные антигены может способствовать вторичным инфекциям. Люди
у больных малярией или трипаносомозом обнаружено подавление иммунитета
к множеству гетерологичных антигенов. Вторичные инфекции часто могут быть
замешан в смерти от африканского трипаносомоза.
Было предложено множество механизмов для объяснения иммуносупрессии.
наблюдается при простейших инфекциях. Наиболее распространенные предлагаемые механизмы: (1)
наличие в инфицированном хозяине паразитов или веществ-хозяев, которые
неспецифически стимулировать рост В-клеток, продуцирующих антитела, а не
стимуляция пролиферации специфических антипаразитарных B-клеток; (2)
пролиферация супрессорных Т-клеток и / или макрофагов, подавляющих иммунную
система выведения регуляторных цитокинов; и (3) продукция паразитом
специфических иммуносупрессоров.
Ссылки
Blackwell JM (ed): Генетика устойчивости к
Бактериальные и паразитарные инфекции. Тейлор и Фрэнсис, Филадельфия, 1988 г.
.- Capron A, Dessaint JP. Молекулярные основы взаимоотношений паразит-хозяин: в сторону
определение защитных антигенов. Иммун Ред.1989; 112: 27. [PubMed: 26 - ]
- Cox FEG, Liew FY. Подмножества Т-клеток и цитокины у паразитарных
инфекции. Паразитол сегодня. 1992; 8: 371. [PubMed: 15463544] Crompton DWT: пищевые взаимодействия между хостами
и паразиты.В: Toft CA, Aeschlimann A, Bolis L (ред.): Parasite-Host
Ассоциация: сосуществование или конфликт. Издательство Оксфордского университета, 1991.- Денис М., Чейди К. Иммунопатология Entamoeba histolytica
инфекции. Паразитол сегодня. 1988; 4: 247. [PubMed: 15463113] - Dyer M, Tait A. Контроль лимфопролиферации с помощью Theilera
anulata . Паразитол сегодня. 1987; 3: 309. [PubMed: 15462874] Englund PT, Sher A (eds): Биология
Паразитизм. Алан Р. Лисс, Нью-Йорк, 1989.- Erard F, LeGros G. Th-2-подобные CD-8 Т-клетки их роль в защите от
Инфекционные заболевания. Паразитол сегодня. 1994; 10: 313. [PubMed: 15275430] - Frenkel JK. Патофизиология токсоплазмоза. Паразитол сегодня. 1988; 4: 273. [PubMed: 15463000]
- Хэдли Т.Дж., Клотц Ф.В., Миллер Л.Х. Поражение эритроцитов малярийными паразитами: клеточные и
молекулярный обзор. Ann Rev Microbiol. 1986; 40: 457. [PubMed: 3535649] Tanner M, Teuscher T, Alonso PL.SPf66 — первая вакцина против малярии. Паразитол сегодня. 1995; 11: 10.
Turner M: Антигенная изменчивость у паразитарных
простейшие. В Birbeck TH, Penn CW (eds): Antigenic Variation in Infectious
Болезни. IRL Press, Оксфорд, Великобритания, 1986.Wakelin D: Иммунитет к паразитам: как животные
Борьба с паразитарными инфекциями. Эдвард Арнольд, Лондон, Великобритания, 1984 год.
Влияние гибели клеток простейших на взаимодействия паразит-хозяин и патогенез | Паразиты и переносчики
Kroemer G, Galluzzi L, Vandenabeele P, Abrams J, Alnemri ES, Baehrecke EH, Blagosklonny MV, El-Deiry WS, Golstein P, Green DR, Hengartner M, Knight RA, Kumar S, Lipton SA, Malorni W, Nuñez G , Питер М.Э., Чопп Дж., Юань Дж., Пьячентини М., Животовский Б., Мелино Г., Номенклатурный комитет по клеточной смерти 2009: Классификация клеточной смерти: рекомендации Номенклатурного комитета по клеточной смерти 2009. Клеточная смерть отличается. 2009, 16: 3-11. 10.1038 / cdd.2008.150.
PubMed Central
CAS
PubMed
Статья
Google Scholar
van Zandbergen G, Luder CG, Heussler V, Duszenko M: Запрограммированная гибель клеток у одноклеточных паразитов: предпосылка для устойчивой инфекции ?. Trends Parasitol. 2010, 26: 477-483. 10.1016 / июл 2010.06.008.
PubMed
Статья
Google Scholar
Bruchhaus I, Roeder T, Rennenberg A, Heussler VT: Простейшие паразиты: запрограммированная смерть клеток как механизм паразитизма. Trends Parasitol. 2007, 23: 376-383. 10.1016 / j.пт.2007.06.004.
CAS
PubMed
Статья
Google Scholar
Хименес-Руис А., Альзате Дж. Ф., Маклеод Е. Т., Людер К. Г., Фазель Н., Херд Н. Маркеры апоптоза у простейших паразитов. Векторы паразитов. 2010, 3: 104-10.1186 / 1756-3305-3-104.
PubMed Central
PubMed
Статья
CAS
Google Scholar
Кубата Б.К., Нагамунэ К., Мураками Н., Меркель П., Кабутуту З., Мартин С.К., Калулу Т.М., Хук М., Йошида М., Охниши-Камеяма М., Киношита Т., Дусенко М., Ураде Y: Kola acuminidins proanthocyanhocyanhocyan. : класс анти-трипаносомных соединений, эффективных против Trypanosoma brucei.Int J Parasitol. 2005, 35: 91-103. 10.1016 / j.ijpara.2004.10.019.
CAS
PubMed
Статья
Google Scholar
Menna-Barreto RF, Salomao K, Dantas AP, Santa-Rita RM, Soares MJ, Barbosa HS, de Castro SL: Различные пути гибели клеток, индуцированные лекарствами в Trypanosoma cruzi: ультраструктурное исследование. Микрон. 2009, 40: 157-168. 10.1016 / j.micron.2008.08.003.
CAS
PubMed
Статья
Google Scholar
Porter H, Gamette MJ, Cortes-Hernandez DG, Jensen JB: Бесполые стадии крови Plasmodium falciparum проявляют признаки вторичного некроза, но не классического апоптоза после воздействия фебрильной температуры (40 C). J Parasitol. 2008, 94: 473-480. 10.1645 / GE-1343.1.
CAS
PubMed
Статья
Google Scholar
Meslin B, Barnadas C, Boni V, Latour C, De Monbrison F, Kaiser K, Picot S: Особенности апоптоза на эритроцитарной стадии Plasmodium falciparum через предполагаемую роль метакаспазоподобного белка PfMCA1.J Infect Dis. 2007, 195: 1852–1859. 10.1086 / 518253.
CAS
PubMed
Статья
Google Scholar
Ньякерига А.М., Перлманн Х., Хагштедт М., Берзиньш К., Трой-Бломберг М., Животовский Б., Перлманн П., Грандиен А: вызванная лекарственными препаратами смерть Plasmodium falciparum на стадиях бесполой крови происходит без типичных признаков апоптоза. . Микробы заражают. 2006, 8: 1560-1568. 10.1016 / j.micinf.2006.01.016.
CAS
PubMed
Статья
Google Scholar
Festjens N, Vanden Berghe T, Vandenabeele P: Некроз, хорошо организованная форма гибели клеток: сигнальные каскады, важные медиаторы и сопутствующий иммунный ответ. Biochim Biophys Acta. 2006, 1757: 1371-1387. 10.1016 / j.bbabio.2006.06.014.
CAS
PubMed
Статья
Google Scholar
Дегтерев А., Хитоми Дж., Гермшайд М., Чэнь И.Л., Коркина О, Тенг Х, Эбботт Д., Куни Г.Д., Юань С., Вагнер Г., Хедрик С.М., Гербер С.А., Луговской А., Юань Дж .: Идентификация киназы RIP1 как специфической клеточной мишени некростатинов.Nat Chem Biol. 2008, 4: 313-321. 10.1038 / nchembio.83.
CAS
PubMed
Статья
Google Scholar
Киль JA: Аутофагия у одноклеточных эукариот. Philos Trans R Soc Lond B Biol Sci. 2010, 365: 819-830. 10.1098 / rstb.2009.0237.
PubMed Central
CAS
PubMed
Статья
Google Scholar
Майури М.С., Зальквар Э., Кимчи А., Кремер Г.: Самопоедание и самоубийство: перекрестные помехи между аутофагией и апоптозом.Обзоры природы. 2007, 8: 741-752.
CAS
PubMed
Статья
Google Scholar
Бера А., Сингх С., Нагарадж Р., Вайдья Т: Индукция гибели аутофагических клеток у Leishmania donovani антимикробными пептидами. Мол Биохим Паразитол. 2003, 127: 23-35. 10.1016 / S0166-6851 (02) 00300-6.
CAS
PubMed
Статья
Google Scholar
Uzcategui NL, Carmona-Gutierrez D, Denninger V, Schoenfeld C, Lang F, Figarella K, Duszenko M: Антипролиферативный эффект дигидроксиацетона на формы кровотока Trypanosoma brucei: прогрессирование клеточного цикла, изменения субклеточной гибели.Противомикробные средства и химиотерапия. 2007, 51: 3960-3968. 10.1128 / AAC.00423-07.
PubMed Central
CAS
PubMed
Статья
Google Scholar
Дельгадо М., Андерсон П., Гарсия-Сальседо Дж. А., Каро М., Гонсалес-Рей Е.: Нейропептиды убивают африканские трипаносомы, воздействуя на внутриклеточные компартменты и вызывая гибель клеток, подобную аутофагии. Смерть клетки отличается. 2009, 16: 406-416. 10.1038 / cdd.2008.161.
CAS
PubMed
Статья
Google Scholar
Санта-Рита Р.М., Лира Р., Барбоса Х.С., Урбина Дж. А., де Кастро С.Л.: Антипролиферативная синергия аналогов лизофосфолипида и кетоконазола против Trypanosoma cruzi (Kinetoplastida: Trypanosomatidae): клеточный и ультраструктурный анализ. J Antimicrob Chemother. 2005, 55: 780-784. 10.1093 / jac / dki087.
CAS
PubMed
Статья
Google Scholar
Menna-Barreto RF, Correa JR, Cascabulho CM, Fernandes MC, Pinto AV, Soares MJ, De Castro SL: Нафтоимидазолы способствуют различным фенотипам смерти у Trypanosoma cruzi.Паразитология. 2009, 136: 499-510. 10.1017 / S003118200
45.
CAS
PubMed
Статья
Google Scholar
Totino PR, Daniel-Ribeiro CT, Corte-Real S, de Fatima Ferreira-da-Cruz M: Plasmodium falciparum: эритроцитарные стадии умирают из-за аутофагической гибели клеток под давлением лекарств. Exp Parasitol. 2008, 118: 478-486. 10.1016 / j.exppara.2007.10.017.
CAS
PubMed
Статья
Google Scholar
Фигарелла К., Равер М., Узкатеги Н.Л., Кубата Б.К., Лаубер К., Мадео Ф., Вессельборг С., Душенко М.: Простагландин D2 вызывает запрограммированную гибель клеток в форме кровотока Trypanosoma brucei. Смерть клетки отличается. 2005, 12: 335-346. 10.1038 / sj.cdd.4401564.
CAS
PubMed
Статья
Google Scholar
Цуда А., Витола У.Х., Охаши К., Онума М.: Экспрессия альтернативной оксидазы ингибирует феномен запрограммированной гибели клеток в форме кровотока Trypanosoma brucei rhodesiense.Parasitol Int. 2005, 54: 243-251. 10.1016 / j.parint.2005.06.007.
CAS
PubMed
Статья
Google Scholar
Велберн С.К., Дейл С., Эллис Д., Бикрофт Р., Пирсон Т.В.: Апоптоз в проциклических Trypanosoma brucei rhodesiense in vitro. Смерть клетки отличается. 1996, 3: 229-236.
CAS
PubMed
Google Scholar
Welburn SC, Модлин I. Борьба с инфекциями Trypanosoma brucei brucei у мухи цеце, Glossina morsitans.Med Vet Entomol. 1997, 11: 286-289. 10.1111 / j.1365-2915.1997.tb00408.x.
CAS
PubMed
Статья
Google Scholar
Vassella E, Reuner B, Yutzy B, Boshart M: Дифференциация африканских трипаносом контролируется механизмом определения плотности, который сигнализирует об остановке клеточного цикла через путь цАМФ. J Cell Sci. 1997, 110 (Pt 21): 2661-2671.
CAS
PubMed
Google Scholar
Figarella K, Uzcategui NL, Beck A, Schoenfeld C, Kubata BK, Lang F, Duszenko M: вызванная простагландином запрограммированная гибель клеток у Trypanosoma brucei связана с окислительным стрессом. Смерть клетки отличается. 2006, 13: 1802-1814. 10.1038 / sj.cdd.4401862.
CAS
PubMed
Статья
Google Scholar
Кубата Б.К., Дусенко М., Кабутуту З., Равер М., Саллиес А., Фудзимори К., Инуи Т., Нозаки Т., Ямасита К., Хории Т., Ураде Ю., Хаяиши О: Идентификация нового простагландина f (2 альфа) синтаза в Trypanosoma brucei.J Exp Med. 2000, 192: 1327-1338. 10.1084 / jem.192.9.1327.
PubMed Central
CAS
PubMed
Статья
Google Scholar
Pentreath VW, Rees K, Owolabi OA, Philip KA, Doua F: Простагландин D2, подавляющий сомногенные Т-лимфоциты, избирательно повышается в спинномозговой жидкости пациентов с запущенной сонной болезнью. Trans R Soc Trop Med Hyg. 1990, 84: 795-799. 10.1016 / 0035-9203 (90)
-С.
CAS
PubMed
Статья
Google Scholar
Welburn SC, Lillico S, Murphy NB: Запрограммированная гибель клеток в проциклической форме Trypanosoma brucei rhodesiense — идентификация дифференциально экспрессируемых генов во время смерти, индуцированной con A. Mem Inst Oswaldo Cruz. 1999, 94: 229-234. 10.1590 / S0074-0276199
CAS
PubMed
Статья
Google Scholar
Welburn SC, Barcinski MA, Williams GT: Запрограммированная гибель клеток у трипаносоматид. Паразитол сегодня. 1997, 13: 22-26.10.1016 / S0169-4758 (96) 10076-4.
CAS
PubMed
Статья
Google Scholar
Дусенко М., Фигарелла К., Маклеод Э.Т., Велберн С.К .: Смерть трипаносомы: эгоистичный альтруизм. Trends Parasitol. 2006, 22: 536-542. 10.1016 / j.pt.2006.08.010.
PubMed
Статья
Google Scholar
Модлин I, Велберн SC: Лектин-опосредованное установление инфекций средней кишки Trypanosoma congolense и Trypanosoma brucei у Glossina morsitans.Троп Мед Паразитол. 1987, 38: 167-170.
CAS
PubMed
Google Scholar
Hu Y, Aksoy S: антимикробный пептид с трипаноцидной активностью, характерный для Glossina morsitans morsitans. Насекомое Biochem Mol Biol. 2005, 35: 105-115. 10.1016 / j.ibmb.2004.10.007.
CAS
PubMed
Статья
Google Scholar
MacLeod ET, Maudlin I., Darby AC, Welburn SC: Антиоксиданты способствуют возникновению трипаносомных инфекций у мухи цеце.Паразитология. 2007, 134: 827-831. 10.1017 / S0031182007002247.
CAS
PubMed
Статья
Google Scholar
Аль-Олайян Э.М., Уильямс Г.Т., Херд Х .: Апоптоз простейших малярии, Plasmodium berghei: возможный механизм ограничения интенсивности инфекции у комаров. Int J Parasitol. 2002, 32: 1133-1143. 10.1016 / S0020-7519 (02) 00087-5.
CAS
PubMed
Статья
Google Scholar
Arambage SC, Grant KM, Pardo I, Ranford-Cartwright L, Hurd H: оокинеты малярии обнаруживают множественные маркеры апоптозоподобной запрограммированной гибели клеток in vitro. Векторы паразитов. 2009, 2: 32-10.1186 / 1756-3305-2-32.
PubMed Central
PubMed
Статья
CAS
Google Scholar
Le Chat L, Sinden RE, Dessens JT: Роль метакаспазы 1 в развитии Plasmodium berghei и апоптозе. Мол Биохим Паразитол. 2007, 153: 41-47.10.1016 / j.molbiopara.2007.01.016.
PubMed Central
CAS
PubMed
Статья
Google Scholar
Pollitt LC, Colegrave N, Khan SM, Sajid M, Reece SE: Исследование эволюции апоптоза у малярийных паразитов: важность экологии. Векторы паразитов. 2010, 3: 105-10.1186 / 1756-3305-3-105.
PubMed Central
PubMed
Статья
Google Scholar
Недельку А.М.: Сравнительная геномика филогенетически разнообразных одноклеточных эукариот дает новое понимание генетической основы эволюции механизма запрограммированной гибели клеток. J Mol Evol. 2009, 68: 256-268. 10.1007 / s00239-009-9201-1.
CAS
PubMed
Статья
Google Scholar
Клемба М., Голдберг Д.Е.: Биологические роли протеаз у паразитических простейших. Анну Рев Биохим. 2002, 71: 275-305. 10.1146 / аннурьев.biochem.71.0
- .145453.
CAS
PubMed
Статья
Google Scholar
Ch’ng JH, Kotturi SR, Chong AG-L, Lear MJ, Tan KS-W: Путь запрограммированной гибели клеток у малярийного паразита Plasmodium falciparum имеет общие черты апоптоза млекопитающих, но опосредуется кланом CA цистеиновые протеазы. Смерть клетки и болезнь. 2010,
Google Scholar
Пико С., Бернод Дж., Бракки В., Чумпитази Б.Ф., Амбруаз-Томас П.: Апоптоз, связанный с чувствительностью к хлорохину малярийного паразита человека Plasmodium falciparum.Trans R Soc Trop Med Hyg. 1997, 91: 590-591. 10.1016 / S0035-9203 (97)
-0.
CAS
PubMed
Статья
Google Scholar
Депонте М., Беккер К.: Plasmodium falciparum — совершают ли убийцы самоубийства ?. Trends Parasitol. 2004, 20: 165-169. 10.1016 / j.pt.2004.01.012.
CAS
PubMed
Статья
Google Scholar
Oakley MS, Kumar S, Anantharaman V, Zheng H, Mahajan B, Haynes JD, Moch JK, Fairhurst R, McCutchan TF, Aravind L. .Заражение иммунной. 2007, 75: 2012-2025. 10.1128 / IAI.01236-06.
PubMed Central
CAS
PubMed
Статья
Google Scholar
Ameisen JC, Idziorek T, Billaut-Mulot O, Loyens M, Tissier JP, Potentier A, Ouaissi A: Апоптоз у одноклеточного эукариота (Trypanosoma cruzi): значение для эволюционного происхождения и роли запрограммированной гибели клеток в контроле клеточной пролиферации, дифференциации и выживания. Смерть клетки отличается.1995, 2: 285-300.
CAS
PubMed
Google Scholar
Zhang J, Andrade ZA, Yu ZX, Andrade SG, Takeda K, Sadirgursky M, Ferrans VJ: Апоптоз на собачьей модели острого шагасического миокардита. J Mol Cell Cardiol. 1999, 31: 581-596. 10.1006 / jmcc.1998.0893.
CAS
PubMed
Статья
Google Scholar
de Souza EM, Araujo-Jorge TC, Bailly C, Lansiaux A, Batista MM, Oliveira GM, Soeiro MN: апоптоз хозяина и паразита после инфекции Trypanosoma cruzi на моделях in vitro и in vivo.Cell Tissue Res. 2003, 314: 223-235. 10.1007 / s00441-003-0782-5.
CAS
PubMed
Статья
Google Scholar
Хименес В., Паредес Р., Соса М.А., Галанти Н.: естественная запрограммированная гибель клеток в эпимастиготах T. cruzi, поддерживаемых в аксенических культурах. Журнал клеточной биохимии. 2008, 105: 688-698. 10.1002 / jcb.21864.
CAS
PubMed
Статья
Google Scholar
De Souza EM, Nefertiti AS, Bailly C, Lansiaux A, Soeiro MN: Дифференциальная апоптозная гибель клеток в амастиготных и трипомастиготных формах из сердечных клеток, инфицированных Trypanosoma cruzi, in vitro. Cell Tissue Res. 2010, 341: 173-180. 10.1007 / s00441-010-0985-5.
CAS
PubMed
Статья
Google Scholar
van Zandbergen G, Solbach W, Laskay T: Инфекция, вызванная апоптозом. Аутоиммунитет. 2007, 40: 349-352. 10.1080 / 08916930701356960.
CAS
PubMed
Статья
Google Scholar
Huynh ML, Fadok VA, Henson PM: Фосфатидилсерин-зависимый прием апоптотических клеток способствует секреции TGF-бета1 и разрешению воспаления. J Clin Invest. 2002, 109: 41-50.
PubMed Central
CAS
PubMed
Статья
Google Scholar
Лаубер К., Блюменталь С.Г., Вайбель М., Вессельборг С.: Очистка апоптотических клеток: избавление от трупов.Mol Cell. 2004, 14: 277-287. 10.1016 / S1097-2765 (04) 00237-0.
CAS
PubMed
Статья
Google Scholar
Волл Р.Е., Херрманн М., Рот Е.А., Стах С., Калден Дж. Р., Гирконтаите I. Иммуносупрессивные эффекты апоптотических клеток. Природа. 1997, 390: 350-351. 10.1038 / 37022.
CAS
PubMed
Статья
Google Scholar
Лаубер К., Бон Э., Кробер С.М., Сяо Ю.Дж., Блюменталь С.Г., Линдеманн Р.К., Марини П., Видиг К., Зобывалский А., Бакш С., Сюй И., Аутенриет И.Б., Шульце-Остхофф К., Белка С., Stuhler G, Wesselborg S: Апоптотические клетки индуцируют миграцию фагоцитов через опосредованное каспазой-3 высвобождение сигнала липидного притяжения.Клетка. 2003, 113: 717-730. 10.1016 / S0092-8674 (03) 00422-7.
CAS
PubMed
Статья
Google Scholar
Фадок В.А., Браттон Д.Л., Фраш С.К., Уорнер М.Л., Хенсон П.М.: Роль фосфатидилсерина в распознавании апоптотических клеток фагоцитами. Смерть клетки отличается. 1998, 5: 551-562. 10.1038 / sj.cdd.4400404.
CAS
PubMed
Статья
Google Scholar
Meagher LC, Savill JS, Baker A, Fuller RW, Haslett C: фагоцитоз апоптотических нейтрофилов не вызывает высвобождение макрофагами тромбоксана B2. J Leukoc Biol. 1992, 52: 269-273.
CAS
PubMed
Google Scholar
Savill JS, Wyllie AH, Henson JE, Walport MJ, Henson PM, Haslett C: Макрофагальный фагоцитоз стареющих нейтрофилов при воспалении. Запрограммированная гибель клеток нейтрофила приводит к его распознаванию макрофагами.J Clin Invest. 1989, 83: 865-875. 10.1172 / JCI113970.
PubMed Central
CAS
PubMed
Статья
Google Scholar
Freire-de-Lima CG, Xiao YQ, Gardai SJ, Bratton DL, Schiemann WP, Henson PM: апоптотические клетки посредством трансформации фактора роста бета координированно индуцируют противовоспалительные и подавляют провоспалительные эйкозаноиды и Синтез NO в мышиных макрофагах. J Biol Chem. 2006, 281: 38376-38384. 10.1074 / jbc.M605146200.
CAS
PubMed
Статья
Google Scholar
Aliberti J, Hieny S, Reis e Sousa C, Serhan CN, Sher A: Липоксин-опосредованное ингибирование продукции IL-12 DC: механизм регуляции микробного иммунитета. Nat Immunol. 2002, 3: 76-82. 10.1038 / ni745.
CAS
PubMed
Статья
Google Scholar
van Zandbergen G, Hermann N, Laufs H, Solbach W., Laskay T. Промастиготы Leishmania выделяют хемотаксический фактор гранулоцитов и вызывают высвобождение интерлейкина-8, но ингибируют продукцию индуцируемого гамма-интерфероном белка 10 нейтрофильными гранулоцитами.Заражение иммунной. 2002, 70: 4177-4184. 10.1128 / IAI.70.8.4177-4184.2002.
PubMed Central
CAS
PubMed
Статья
Google Scholar
van Zandbergen G, Bollinger A, Wenzel A, Kamhawi S, Voll R, Klinger M, Muller A, Holscher C, Herrmann M, Sacks D, Solbach W., Laskay T. Развитие болезни лейшмании зависит от наличия апоптотических промастигот в вирулентном инокуляте. Proc Natl Acad Sci USA. 2006, 103: 13837-13842.10.1073 / pnas.0600843103.
PubMed Central
CAS
PubMed
Статья
Google Scholar
Wanderley JL, Pinto da Silva LH, Deolindo P, Soong L, Borges VM, Prates DB, de Souza AP, Barral A, Balanco JM, do Nascimento MT, Saraiva EM, Barcinski MA: Сотрудничество между апоптозом и жизнеспособные метациклики усиливают патогенез лейшманиоза. PLoS One. 2009, 4: e5733-10.1371 / journal.pone.0005733.
PubMed Central
PubMed
Статья
CAS
Google Scholar
ван Зандберген Г., Клингер М., Мюллер А., Данненберг С., Геберт А., Сольбах В., Ласкей Т. Передний край: нейтрофильные гранулоциты служат вектором проникновения лейшмании в макрофаги. J Immunol. 2004, 173: 6521-6525.
CAS
PubMed
Статья
Google Scholar
Риттер У., Фришкнехт Ф., ван Зандберген Г: Важны ли нейтрофилы клетками-хозяевами для паразитов Leishmania ?. Trends Parasitol. 2009, 25: 505-510. 10.1016 / j.пт.2009.08.003.
CAS
PubMed
Статья
Google Scholar
Peters NC, Egen JG, Secundino N, Debrabant A, Kimblin N, Kamhawi S, Lawyer P, Fay MP, Germain RN, Sacks D: визуализация in vivo показывает важную роль нейтрофилов в лейшманиозе, передающемся через песок. летит. Наука. 2008, 321: 970-974. 10.1126 / science.1159194.
PubMed Central
CAS
PubMed
Статья
Google Scholar
Wanderley JL, Moreira ME, Benjamin A, Bonomo AC, Barcinski MA: Мимикрия апоптотических клеток путем экспонирования фосфатидилсерина участвует в создании амастигот Leishmania (L) amazonensis у млекопитающих-хозяев. J Immunol. 2006, 176: 1834-1839.
CAS
PubMed
Статья
Google Scholar
Даматта Р.А., Сибра С.Х., Деолиндо П., Арнхольдт А.С., Манхес Л., Гольденберг С., де Соуза В.: Trypanosoma cruzi раскрывает фосфатидилсерин как механизм уклонения.FEMS Microbiol Lett. 2007, 266: 29-33. 10.1111 / j.1574-6968.2006.00495.x.
CAS
PubMed
Статья
Google Scholar
Lopes MF, da Veiga VF, Santos AR, Fonseca ME, DosReis GA: Гибель CD4 + Т-клеток, вызванная активацией, в результате апоптоза при экспериментальной болезни Шагаса. J Immunol. 1995, 154: 744-752.
CAS
PubMed
Google Scholar
Freire-de-Lima CG, Nascimento DO, Soares MB, Bozza PT, Castro-Faria-Neto HC, de Mello FG, DosReis GA, Lopes MF: Поглощение апоптотических клеток стимулирует рост патогенных трипаносом в макрофагах.Природа. 2000, 403: 199-203. 10.1038 / 35003208.
CAS
PubMed
Статья
Google Scholar
Ribeiro-Gomes FL, Otero AC, Gomes NA, Moniz-De-Souza MC, Cysne-Finkelstein L, Arnholdt AC, Calich VL, Coutinho SG, Lopes MF, DosReis GA: Взаимодействие макрофагов с нейтрофилами регулирует лейшманию. серьезная инфекция. J Immunol. 2004, 172: 4454-4462.
CAS
PubMed
Статья
Google Scholar
Мамани-Мацуда М., Рамберт Дж., Мальви Д., Лежоли-Буассо Н., Даулуэд С., Тиолат Д., Коув С., Куртуа П., Винсендо П., Мосалаи М.Д .: Кверцетин индуцирует апоптоз трипаносомы макропротеина человека и снижает провоспалительную реакцию человека. . Противомикробные агенты Chemother. 2004, 48: 924-929. 10.1128 / AAC.48.3.924-929.2004.
PubMed Central
CAS
PubMed
Статья
Google Scholar
Seabra SH, de Souza W., Damatta RA: Toxoplasma gondii обнажает фосфатидилсерин, вызывая аутокринный эффект TGF-бета1, управляющий уклонением макрофагов.Biochem Biophys Res Commun. 2004, 324: 744-752. 10.1016 / j.bbrc.2004.09.114.
CAS
PubMed
Статья
Google Scholar
Ван Ю., Вайс Л. М., Орлофски А. Аутофагия клетки-хозяина индуцируется Toxoplasma gondii и способствует росту паразитов. J Biol Chem. 2009, 284: 1694-1701. 10.1074 / jbc.M8078
.
PubMed Central
CAS
PubMed
Статья
Google Scholar
van Zandbergen G, Luder CG, Heussler V, Duszenko M: Запрограммированная гибель клеток у одноклеточных паразитов: предпосылка для устойчивой инфекции ?. Trends Parasitol. 2010, 26: 477-83. 10.1016 / июл 2010.06.008.
PubMed
Статья
Google Scholar
Sturm A, Amino R, van de Sand C, Regen T, Retzlaff S, Rennenberg A, Krueger A, Pollok JM, Menard R, Heussler VT: Манипуляции с гепатоцитами хозяина малярийным паразитом для доставки в печень синусоиды.Наука. 2006, 313: 1287-1290. 10.1126 / science.1129720.
CAS
PubMed
Статья
Google Scholar
Бестейро С., Уильямс Р.А., Моррисон Л.С., Кумбс Г.Х., Моттрам Дж.С.: Сортировка эндосом и аутофагия важны для дифференциации и вирулентности Leishmania major. J Biol Chem. 2006, 281: 11384-11396. 10.1074 / jbc.M512307200.
CAS
PubMed
Статья
Google Scholar
Williams RA, Tetley L, Mottram JC, Coombs GH: Цистеинпептидазы CPA и CPB жизненно важны для аутофагии и дифференциации Leishmania mexicana. Mol Microbiol. 2006, 61: 655-674. 10.1111 / j.1365-2958.2006.05274.x.
CAS
PubMed
Статья
Google Scholar
Уильямс Р.А., Вудс К.Л., Джулиано Л., Моттрам Дж. К., Кумбс Г. Х .: Характеристика необычных семейств ATG8-подобных белков и ATG12 у простейшего паразита Leishmania major.Аутофагия. 2009, 5: 159-172. 10.4161 / авто.5.2.7328.
PubMed Central
CAS
PubMed
Статья
Google Scholar
Баккер Б.М., Вестерхофф Х.В., Михельс П.А.: Регулирование и контроль компартментализованного гликолиза в кровотоке формы Trypanosoma brucei. Журнал биоэнергетики и биомембран. 1995, 27: 513-525. 10.1007 / BF02110191.
CAS
PubMed
Статья
Google Scholar
Hannaert V, Michels PA: Структура, функция и биогенез гликозом в кинетопластидах. Журнал биоэнергетики и биомембран. 1994, 26: 205-212. 10.1007 / BF00763069.
CAS
PubMed
Статья
Google Scholar
Michels PA, Bringaud F, Herman M, Hannaert V: Метаболические функции гликозом у трипаносоматид. Biochimica et biophysica acta. 2006, 1763: 1463-1477.
CAS
PubMed
Статья
Google Scholar
Герман М., Перес-Морга Д., Штикцелле Н., Михельс П.А.: Оборот гликозом во время дифференциации жизненного цикла Trypanosoma brucei. Аутофагия. 2008, 4: 294-308.
CAS
PubMed
Статья
Google Scholar
Ригден Д. Д., Герман М., Гиллис С., Михельс П. А.: Последствия геномного поиска генов, связанных с аутофагией, у трипаносоматид. Biochem Soc Trans. 2005, 33: 972-974. 10.1042 / BST20050972.
CAS
PubMed
Статья
Google Scholar
Фигейредо Р.К., Роза Д.С., Соарес М.Дж.: Дифференциация эпимастигот Trypanosoma cruzi: метациклогенез и адгезия к субстрату запускаются стрессом, связанным с питанием. Журнал паразитологии. 2000, 86: 1213-1218. 10.2307 / 3285002.
CAS
PubMed
Статья
Google Scholar
Альварес В.Е., Косек Г., Сант’Анна К., Тюрк В., Каззуло Дж. Дж., Тюрк Б. Аутофагия участвует в реакции на пищевой стресс и дифференциации у Trypanosoma cruzi.Журнал биологической химии. 2008, 283: 3454-3464. 10.1074 / jbc.M708474200.
CAS
PubMed
Статья
Google Scholar
Коллиен А.Х., Шауб Г.А.: Развитие Trypanosoma cruzi у triatominae. Паразитология сегодня (личное ред.). 2000, 16: 381-387. 10.1016 / S0169-4758 (00) 01724-5.
CAS
Статья
Google Scholar
Соарес М.Дж., Соуто-Падрон Т., Боналдо М.К., Гольденберг С., де Соуза В.: стереологическое исследование процесса дифференциации у Trypanosoma cruzi.Паразитологические исследования. 1989, 75: 522-527. 10.1007 / BF00931160.
CAS
PubMed
Статья
Google Scholar
Деннингер В., Купманн Р., Мухаммад К., Барт Т., Бассарак Б., Шонфельд С., Килунга Б.К., Душенко М: Кинетопластида: модельные организмы для простых аутофагических путей ?. Методы Энзимол. 2008, 451: 373-408. полный текст.
CAS
PubMed
Статья
Google Scholar
Barquilla A, Crespo JL, Navarro M: Рапамицин подавляет рост трипаносомных клеток, предотвращая образование комплекса 2 TOR. Proc Natl Acad Sci USA. 2008, 105: 14579-14584. 10.1073 / pnas.0802668105.
PubMed Central
CAS
PubMed
Статья
Google Scholar
Scherz-Shouval R, Shvets E, Fass E, Shorer H, Gil L, Elazar Z: активные формы кислорода необходимы для аутофагии и специфически регулируют активность Atg4.Журнал EMBO. 2007, 26: 1749-1760. 10.1038 / sj.emboj.7601623.
PubMed Central
CAS
PubMed
Статья
Google Scholar
Uzcategui NL, Denninger V, Merkel P, Schoenfeld C, Figarella K, Duszenko M: Аутофагия, индуцированная дигидроксиацетоном в африканских трипаносомах. Аутофагия. 2007, 3: 626-629.
CAS
PubMed
Статья
Google Scholar
Dantas AP, Salomao K, Barbosa HS, De Castro SL: Эффект болгарского прополиса против Trypanosoma cruzi и во время его взаимодействия с клетками-хозяевами. Memorias do Instituto Oswaldo Cruz. 2006, 101: 207-211.
PubMed
Статья
Google Scholar
Cunha-e-Silva NL, Atella GC, Porto-Carreiro IA, Morgado-Diaz JA, Pereira MG, De Souza W: Выделение и характеристика фракции резервосом из Trypanosoma cruzi.FEMS Microbiol Lett. 2002, 214: 7-12. 10.1111 / j.1574-6968.2002.tb11317.x.
CAS
PubMed
Статья
Google Scholar
Menna-Barreto RF, Laranja GA, Silva MC, Coelho MG, Paes MC, Oliveira MM, de Castro SL: Масло семян Pterodon pubescens против Trypanosoma cruzi: геранилгераниол в качестве основного биологически активного компонента. Паразитологические исследования. 2008, 103: 111-117. 10.1007 / s00436-008-0937-0.
CAS
PubMed
Статья
Google Scholar
Assuncao Guimaraes C, Linden R: Запрограммированная гибель клеток. Апоптоз и альтернативные стили смерти. Европейский журнал биохимии / FEBS. 2004, 271: 1638-1650. 10.1111 / j.1432-1033.2004.04084.x.
PubMed
Статья
CAS
Google Scholar
Эль-Сайед Н.М., Майлер П.Дж., Бартоломеу Д.К., Нильссон Д., Аггарвал Г., Тран А.Н., Гедин Е., Уорти Е.А., Делчер А.Л., Бландин Г.Последовательность генома Trypanosoma cruzi, этиологического агента болезни Шагаса. .Наука (Нью-Йорк, Нью-Йорк). 2005, 309: 409-415.
CAS
Статья
Google Scholar
Герман М., Гиллис С., Михельс П.А., Ригден Д.Д.: Аутофагия и родственные процессы у трипаносоматид: выводы из геномного и биоинформатического анализа. Аутофагия. 2006, 2: 107-118.
CAS
PubMed
Статья
Google Scholar
Li Q, Zhou Y, Yao C, Ma X, Wang L, Xu W, Wang Z, Qiao Z: апоптоз, вызванный гелданамицином, ингибитором Hsp90, у Leishmania donovani на стадии трансформации промастигот в амастиготу.Parasitol Res. 2009, 105: 1539-1548. 10.1007 / s00436-009-1582-г.
PubMed
Статья
Google Scholar
Альзате Дж. Ф., Ариас А. А., Морено-Матеос Д., Альварес-Барриентос А., Хименес-Руис А. Митохондриальный супероксид опосредует апоптозную смерть, вызванную нагревом, у Leishmania infantum. Мол Биохим Паразитол. 2007, 152: 192-202. 10.1016 / j.molbiopara.2007.01.006.
CAS
PubMed
Статья
Google Scholar
Райна П., Каур С.: Дифференциация, вызванная хроническим тепловым шоком, вызывает апоптоз у Leishmania donovani. Mol Cell Biochem. 2006, 289: 83-90. 10.1007 / s11010-006-9151-5.
CAS
PubMed
Статья
Google Scholar
Альзате Дж. Ф., Альварес-Барриентос А., Гонсалес В. М., Хименес-Руис А.: Вызванная нагреванием запрограммированная смерть клеток у Leishmania infantum восстанавливается экспрессией Bcl-X (L). Апоптоз. 2006, 11: 161-171. 10.1007 / s10495-006-4570-z.
CAS
PubMed
Статья
Google Scholar
Морейра М.Э., Дель Портильо Х.А., Мильдер Р.В., Баланко Дж.М., Барчински М.А.: Индукция апоптоза при тепловом шоке в промастиготах одноклеточного организма Leishmania (Leishmania) amazonensis. J. Cell Physiol. 1996, 167: 305-313. 10.1002 / (SICI) 1097-4652 (199605) 167: 2 <305 :: AID-JCP15> 3.0.CO; 2-6.
CAS
PubMed
Статья
Google Scholar
Микроскопические организмы — Австралийская антарктическая программа
Микроскопические организмы — это крошечные формы жизни, часто состоящие из одной клетки.Они очень чувствительны к изменениям.
Микроскопические организмы жизненно важны в пищевой цепи и для здоровья нашей планеты. Они составляют основу морской пищевой сети и прямо или косвенно служат пищей для всего остального в открытом море.
Микроскопические организмы также играют важную роль в поддержании атмосферы Земли. Они помогают удалять углекислый газ и выделяют химические вещества, которые помогают формировать облака. Ученые изучают микроскопические организмы в Антарктике, чтобы лучше понять атмосферные изменения и истощение озонового слоя.
Четыре основных типа микроорганизмов в океане:
- Водоросли — это одноклеточные растения, также известные как фитопланктон (от греческого означает дрейфующие растения).
- Простейшие — это одноклеточные животные, также известные как зоопланктон (от греческого — дрейфующие животные).
- Бактерии — самые многочисленные организмы на Земле.
- Вирусы — самые распространенные биологические агенты в морской воде.Они заражают фитопланктон, простейшие и бактерии и могут играть важную роль в контроле их численности и состава.
«Протисты» — общий термин для одноклеточных организмов, включая фитопланктон и простейшие.
Размер
Бактерии — это мельчайшие микроорганизмы размером от 0,0001 мм до 0,001 мм. Фитопланктон и простейшие колеблются от примерно 0,001 мм до примерно 0,25 мм. Самый большой фитопланктон и простейшие можно увидеть невооруженным глазом, но большинство из них можно увидеть только под микроскопом.
Изобилие
Эти организмы могут быть крошечными, но они присутствуют в огромных количествах.
В каждом литре морской воды содержится:
- от 1 до 4 миллиардов вирусов
- около 1 млрд бактерий
- около 1 млн фитопланктона
- около полумиллиона простейших
Когда вес и количество этих организмов умножаются вместе, микроорганизмы представляют собой самую большую биомассу на Земле.
Тип
Фитопланктон: морское пастбище
Фитопланктон — это растения.Они получают энергию для роста от солнечного света в процессе фотосинтеза.
В Южном океане обитает около 400 видов фитопланктона. Различные виды различаются по форме и размеру или по фотосинтетическим пигментам. Их также можно отличить по тому, заключены ли они клеточной стенкой, и если да, то из чего эта клеточная стенка сделана. Многие также могут быть идентифицированы по расположению крошечных чешуек и шипов, покрывающих их поверхность.Эти чешуйки настолько малы, что детали их формы и рисунка можно увидеть только с помощью электронного микроскопа с большим увеличением. Очень немногие фитопланктон ядовиты.
Фитопланктон можно рассматривать как морское пастбище. Они обеспечивают пищу и энергию для пищевой сети Южного океана. Большую часть года антарктические моря покрыты льдом. Это, в сочетании с низким углом наклона солнца и короткими днями, ограничивает свет, доступный для фотосинтеза. В результате количество многих видов сокращается в зимние месяцы и снова цветет весной и летом, когда условия освещения более благоприятны.
В верхних слоях океана фитопланктон сопровождается одноклеточными животными, называемыми простейшими. Простейшие питаются бактериями. И фитопланктон, и простейшие поедаются более крупными животными Южного океана, включая криль.
Простейшие
Простейшие встречаются в самых разных средах обитания, включая море, пресную воду, почву и тела других организмов. Фактически, везде, где есть влага, обычно встречаются простейшие. Некоторые виды ограничены холодной водой и встречаются только в Южном океане.
Эти крошечные животные обладают всеми функциями более крупных существ: они поглощают пищу, выделяют отходы, размножаются и общаются. Они питаются непосредственно фитопланктоном, бактериями и другими простейшими. Их дыхание высвобождает большую часть углекислого газа, содержащегося в фитопланктоне. Однако они также помогают удалять углекислый газ из атмосферы, превращая свою микроскопическую пищу в собственную клеточную массу. Примечательно, что такое маленькое отдельное устройство может выполнять все действия, выполняемые более крупными животными.Некоторые простейшие могут выйти из неблагоприятных условий, впадая в спячку.
Бактерии
Бактерии были обнаружены везде, где может существовать жизнь. Они встречаются в глубинах океана и в ледяных покровах на полюсах. Некоторые из них были извлечены и выросли из льда и снега, выпавших много веков назад в Антарктике. Наиболее распространенные виды бактерий имеют размер от 0,0005 мм до 0,005 мм. Это означает, что ряд из 100 самых крупных бактерий достигнет точки в конце этого предложения.
Большинство бактерий являются потребителями. Очень незначительное меньшинство может вызывать болезни у других живых существ, но подавляющее большинство питается отходами или мертвыми организмами. Это организмы-разлагатели. Они имеют жизненно важное значение для жизни на Земле, поскольку распространяют элементы, из которых состоят живые существа.
Несколько бактерий являются продуцентами. Некоторые из них похожи на растения в том, что они производят энергию путем фотосинтеза. Другие бактерии используют химические вещества, такие как сера, железо, газообразный водород или соединения азота, в процессе, называемом хемосинтезом.
Вирусы
Вирусы — самые распространенные биологические агенты в морской воде. Концентрации в антарктических водах колеблются от 1 до 4 миллионов частиц на мл. Они заражают фитопланктон, простейшие и бактерии и могут быть причиной до 50% смертей морских бактерий. Разрывные клетки выпускают свое содержимое в воду, где они способствуют росту бактерий.