
Содержание
Гомологический ряд — это… Что такое Гомологический ряд?
Рис. 1 — Температуры плавления (синий) и кипения (красный) в гомологическом ряду n-алканов C1…C14.
Рис. 2 — Температуры плавления (синий) и кипения (фиолетовый) в гомологическом ряду алифатических карбоновых кислот C1…C8.
Гомологический ряд — ряд химических соединений одного структурного типа (например, алканы или алифатические спирты — спирты жирного ряда), отличающихся друг от друга по составу на определенное число повторяющихся структурных единиц — так называемую «гомологическую разность». Чаще всего это метиленовые звенья: …—СН2—… Простейший пример гомологического ряда — низшие гомологи алканов (общая формула СnH2n+2): метан CH4, этан C2H6, пропан С3H8 и т. д.
Гомология и структура соединений
В основе понятия гомологии в органической химии лежит фундаментальное положение о том, что химические и физические свойства вещества определяются структурой его молекул: эти свойства определяются как функциональными группами соединения (гидроксил спиртов, карбоксильная группа карбоновых кислот, арильная группа ароматических соединений и т. п.), так и его углеродным скелетом.
п.), так и его углеродным скелетом.
Сам комплекс химических свойств и, соответственно, принадлежность соединения к определённому классу, определяется именно функциональными группами (так, наличие карбоксильной группы определяет проявление соединением кислотных свойств и его принадлежность к классу карбоновых кислот), но на степень проявления химических свойств (например, реакционная способность и константа диссоциации) или физические свойства (температуры кипения и плавления, показатель преломления и т. п.) влияет и углеродный скелет молекулы (см. Рис. 1).
В случае подобия углеродных скелетов соединений, то есть отсутствия изомерии, формулу гомологичных соединений можно записать как X—(СН2)n—Y, соединения с различным числом n метиленовых звеньев являются гомологами и принадлежат к одному классу соединений (например, H—(СН2)n—COOH — алифатические карбоновые кислоты). Таким образом, соединения-гомологи принадлежат к одному классу соединений, и свойства ближайших гомологов наиболее близки.
В гомологических рядах наблюдается закономерное изменение свойств от младших членов ряда к старшим, однако такая закономерность может нарушаться, в первую очередь, в начале ряда, из-за образования водородных связей при наличии функциональных групп, способных к их образованию (см. Рис. 2, температуры плавления).
Интересные факты
При исследовании параллелизмов в явлениях наследственной изменчивости, Н. И. Вавиловым, по аналогии с гомологическими рядами органических соединений, было введено понятие Гомологические ряды в наследственной изменчивости.
См. также
Урок химии на тему «Алканы. Гомологический ряд. Номенклатура алканов»
Тип урока: комбинированный
Цели урока:
- Изучить гомологический ряд и строение
веществ ряда алканов. - Научиться давать название веществам,
принадлежащим к ряду алканов.
Оснащение урока:
- Компьютерная презентация (Приложение
1), компьютер, проектор.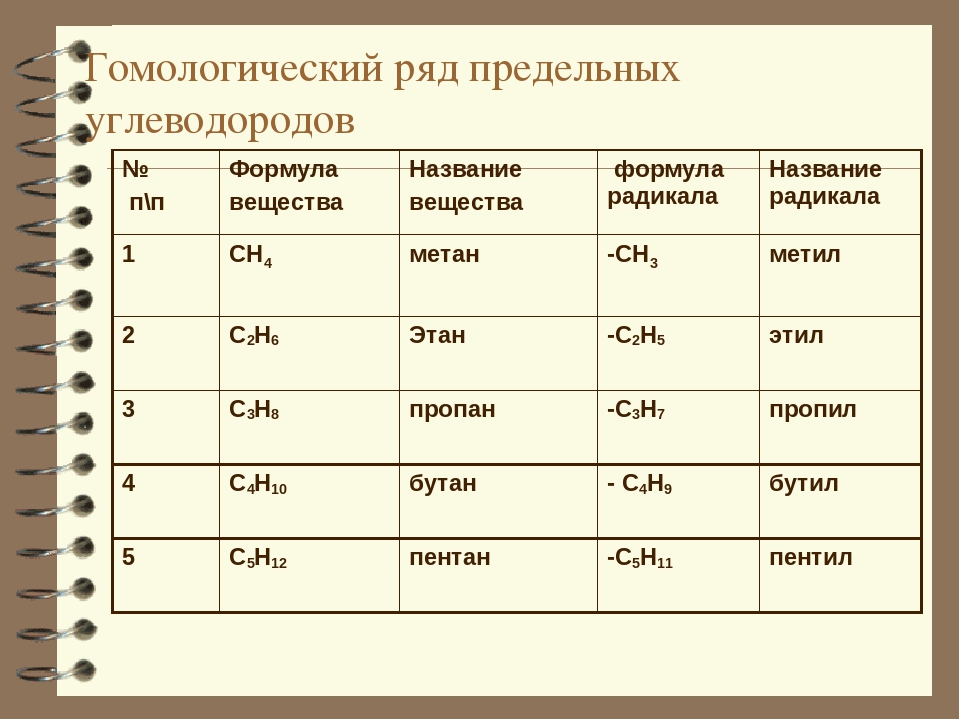
- Раздаточный материал –
- листочки с таблицей «Классификация и
характеристика углеводородов» (Приложение
2), - листочки с таблицей «Гомологический ряд
алканов» (Приложение 3), - листочки «Алгоритм определения названия
органического вещества» (Приложение
4) (на каждом листочке с обратной
стороны прикреплен двусторонний скотч)
- листочки с таблицей «Классификация и
- Карточки с заданиями для парной работы (Приложение 5)
- на нахождение молекулярной формулы алканов
- определение названия алканов
ХОД УРОКА
I. Фронтальная беседа
Вопросы для повторения:
- Какой химический элемент является основой всех
органических веществ? - Что изучает органическая химия?
- Кто из ученых ввел понятие «органическая
химия»? - Дайте определение валентности.

- Какова валентность углерода во всех
органических веществах? - Назовите валентности водорода, галогенов,
кислорода и азота в органических веществах - Какое количество органических веществ
известно? Чем можно объяснить такое
многообразие? - Какой ученый создал теорию строения
органических веществ и тем самым помог их
систематизации? - Что показывают следующие записи формул веществ,
например: - Какие вещества называются гомологами?
II. Проверка выполнения домашнего задания
К доске выходит учащийся и выполняет задание по
написанию возможных изомеров веществ с
молекулярной формулой С6H14.
(Во время выполнения учащимся задания на
доске, остальные учащиеся обмениваются
тетрадями и осуществляют взаимопроверку
домашнего задания, выставляют оценки и
расписываются. )
)
III. Объяснение нового материала
Учитель объявляет тему и цели нового урока:
(слайды № 1,2)
3.1. Рассказ учителя:
На данный момент известно около 60 миллионов
органических веществ. Как
разобраться в таком количестве? Это удобно
делать, когда все вещества распределены на
определенные группы по сходным признакам.
Общепринятая классификация органических
соединений основана именно на теории строения
А.М. Бутлерова.
В любом органическом веществе есть атомы
углерода, которые соединяясь друг с другом,
образуют основу («скелет») и для правильной
структуры вещества и сохранения
четырехвалентности углерода, остальные связи
закрывают атомы водорода.
3.2. Объяснение понятия «Углеводороды»
Учитель задает вопрос: «Как называется группа
веществ, состоящих из улерода и водорода?»
Учащиеся делают вывод «Углеводороды».
Записывают понятие в тетрадь.
Возникает вопрос: какое отношение понятие
«углеводороды» имеет к теме урока?
3.3. Объяснение понятия «Алканы»
Учитель предлагает учащимся найти среди
листочков, лежащих на столе, листок (Приложение
2) «Классификация и характеристика
углеводородов». Исходя из этой таблицы, можно
ответить на поставленный вопрос.
«Алканы» – это класс органических веществ,
относящийся к группе углеводородов. Учитель
предлагает на чистый лист своей тетради
прикрепить при помощи двустороннего скотча Приложение 2. (По мере
изучения различных классов углеводородов
таблица будет заполняться. На данном уроке
заполняется только один столбик, касающийся
алканов.)
При объяснении понятия алканы, учитель
показывает структурные формулы этана, пропана,
бутана и просит учащихся подумать в чем
сходство строения всех этих веществ? (Слайд № 3)
Ученики делают вывод, о том, что, во всех этих
веществах встречаются только простые одинарные
связи. На основании этого вывода в тетрадь
На основании этого вывода в тетрадь
записывается определение «Алканы» или
предельные углеводороды. Также заполняется в
таблице «Классификация и характеристика
углеводородов» пункт «отличительный
признак».
3.4. Изучение гомологического ряда алканов
Приложение 3
«Гомологический ряд алканов» вклеивается в
тетрадь и заполняется совместно с учителем.
(Слайд № 4).
Учитель обращает внимание учащихся, что
порядок расположения веществ зависит от
количества атомов углерода в молекуле и, чтобы
легче было запомнить каким по счету идет то или
иное вещество, можно загибать пальцы на руках
(один палец – один атом углерода). В таблице
заполняются пункты «название вещества» и
«молекулярная формула», а затем все вместе хором
повторяют название всех записанных веществ.
Учитель обращает внимание, что все вещества
заканчиваются на суффикс -ан, так же
как и название класса «алканы».
При заполнении таблицы учитель просит обратить
внимание учащихся на то, чем отличается
формула каждого последующего вещества от
предыдущего
Ученики делают вывод, что все вещества
отличаются друг от друга на группу атомов –СН2.
Учитель на основе вывода дает понятие
гомологов и гомологического ряда. Ученики
записывают в тетрадь понятие «гомологи» (Слайд №
5)
Учитель обращает внимание на то, что количество
углерода и водорода в формулах алканов можно
определить, зная порядковый номер вещества в
гомологическом ряду
(Учитель еще раз напоминает, что для лучшего
заполнения можно использовать пальцы рук) так
как есть общая формула алканов и записывается
она СnH2n+2 (слайд № 6), где маленькая
буква «n» – обозначает количество атомов
углерода в молекуле. Учитель на примере
показывает, каким образом сосчитать количество
атомов водорода, зная количество атомов углерода
в молекуле.
3.5. Выполнение парного задания на нахождение
молекулярной формулы алканов по общей формуле. (Приложение 4)
3.6. Изучение номенклатуры алканов.
В таблице «Гомологический ряд алканов»
(слайд № 4) заполняется пункт «структурная
формула вещества». Первые четыре учитель
заполняет сам, а следующие шесть заполняет
ученик, приглашенный к доске.
Учитель делает вывод о том, что все вещества,
имеющие прямолинейное строение именно так и
будут называться, как записано в таблице.
Для определения названия органических
веществ разветвленного строения существует
международная номенклатуре IUPAK (International Union of Pure and
Applied Chemistry) (или в русской транскрипции – ИЮПАК)
(слайд № 7), такая номенклатура позволяет
понимать ученым всего мира друг друга. Эта
система понятна и лаконична, главное знать
название первых представителей ряда алканов.
Учащиеся прикрепляют листочек «Алгоритм
определения названия органических веществ» в
тетрадь. (Слайд № 8)
(Слайд № 8)
Идет прочтение каждого пункта, после которого
учитель показывает на примере как правильно,
следуя алгоритму дать название веществу (на
примере 2-метилбутана, 2,2-диметилбутана, 2-метил,
3-этилгексана).
При этом учитель обращает внимание на понятие
«радикал» и направляет учеников к таблице № 2
«Гомологический ряд алканов», где остается
незаполненным этот пункт. Ученики заполняют
его, вписывая названия первых четырех радикалов
3.7.Работа в парах. Выполнение задания по
карточкам. (Приложение 5)
IV. Повторение пройденного материала
Фронтальный опрос
Вопросы:
- Какие вещества называют углеводородами?
- Какие классы органических веществ относятся к
углеводородам? - Какие органические вещества называются
алканами? - Общая формула алканов?
- Как определить по этой формуле количество
атомов водорода в молекуле алкана? - Как дать название веществу нелинейного
строения?
V. Подведение итогов урока. Выставление
Подведение итогов урока. Выставление
оценок за урок
VI. Выдача домашнего задания
Список литературы
- Габриелян О.С. Химия для профессий и
специальностей технического профиля/ О.С.
Габриелян, И.Г. Остроумов. – М.: ОИЦ «Академия», 2010
Ароматические углеводороды — Что такое Ароматические углеводороды?
Ароматические углеводороды (арены) – это углеводороды, молекулы которых содержат одно или несколько бензольных колец.
Ароматические углеводороды (арены) — класс углеводородов, содержащих бензольные ядра, которые могут быть конденсированными и иметь насыщенные боковые цепи.
К наиболее важным аренам относятся бензол (С6Н6) и его гомологи.
Это углеводороды, молекулы которых содержат 1 или несколько бензольных колец, и другие углеводородные группы (алифатические, нафтеновые, полициклические).
Простейшим представителем ароматических углеводородов является бензол, молекулярная формула которого С6Н6.
Установлено, что все атомы углерода в молекуле бензола лежат в одной плоскости, образуя правильный 6-угольник .
Каждый атом углерода связан с одним атомом водорода.
Длины всех связей углерод-углерод одинаковы и составляют 0,139 нм.
Для передачи строение молекулы бензола используют формулы Кекуле, которые предложил в 1865 г. немецкий химик Август Кекуле.
Важными аренами являются и гомологи бензола (толуол С6Н5СНз, ксилолы С6Н4(СНз)2, дурол, мезитилен, этилбензол), кумол, нафталин C10H8, антрацен С14Н10 и их производные.
Ароматические углеводороды — исходное сырье для промышленного получения кетонов, альдегидов и кислот ароматического ряда, и других веществ.
Исторически название «ароматические углеводороды» сложилось потому, что многие производные бензола, которые первыми были выделены из природных источников, обладали приятным запахом.
В настоящее время под понятием «ароматичность» подразумевают, прежде всего, особый характер реакционной способности веществ, обусловленный, в свою очередь, особенностями строения молекул этих соединений.
В чем же состоят эти особенности?
В соответствии с молекулярной формулой С6Н6 бензол является ненасыщенным соединением, и можно ожидать, что для него были бы характерны типичные для алкенов реакции присоединения.
Однако в условиях, в которых алкены быстро вступают в реакции присоединения, бензол не реагирует или реагирует медленно.
Бензол не дает и характерных качественных реакций, свойственных непредельным углеводородам: он не обесцвечивает бромную воду и водный раствор перманганата калия.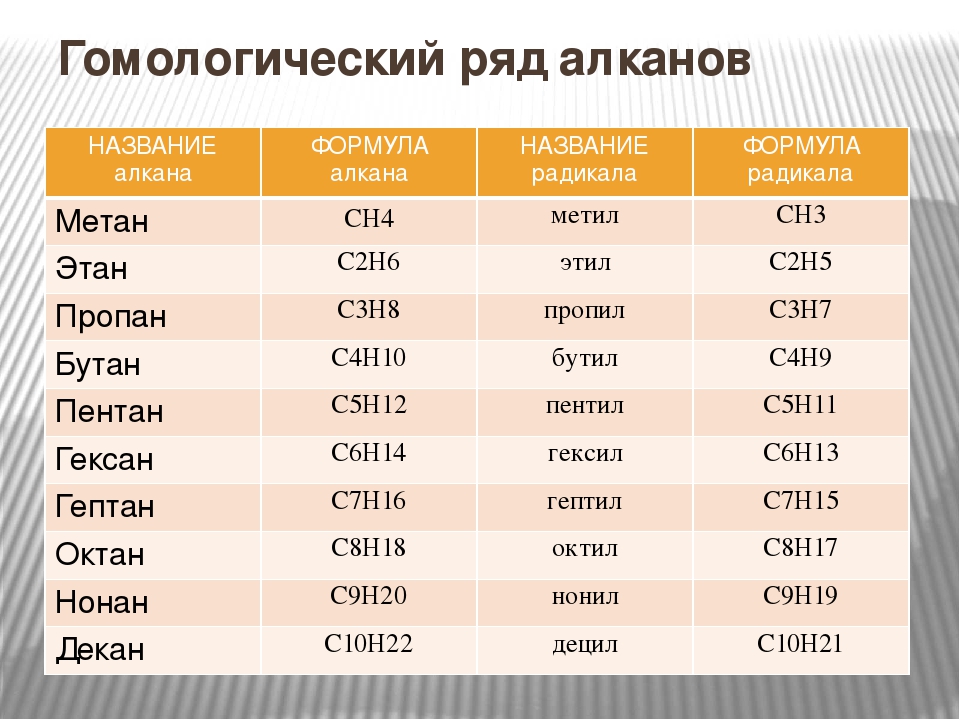
Такой характер реакционной способности объясняется наличием в ароматическом кольце сопряженной системы — единого «пи»-электронного облака.
Гомологический ряд
CH4 —
метан
C2H6 —
этан
C3H8 —
пропан
C4H10 —
бутан
C5H12 —
пентан
C6H14 —
гексан
C7H16 —
гептан
C8H18 —
октан
C9H20 —
нонан
C10H22 —
декан …
Гомологический
ряд характеризуется общей формулой, по
которой можно определить формулу любого
вещества данного ряда. Для предельных
углеводородов —
эта формула
CnH2n+2
C
увеличением молекулярной массы вещества
(увеличивается количество атомов
углерода и водорода в молекуле) происходит
увеличение температуры кипения этого
вещества. Поэтому следует отметить, что
Поэтому следует отметить, что
вещества с CH4 по
C4H10 —
газы, с C5H12 по
C15H32 —
жидкости, далее — это твёрдые вещества!
7.5
Какие органические соединения называются
изомерами, какие разновидности структурной
и пространственной изомерии существуют.
Изомерия —
явление, заключающееся в существовании
химических соединений — изомеров,
— одинаковых по атомному составу и
молекулярной массе, но различающихся
по строению или расположению атомов в
пространстве и, вследствие этого, по
свойствам.
Структурная
изомерия — результат различий в
химическом строени.
Изомерия углеродной цепи (углеродного скелета)[
Изомерия
углеродного скелета, обусловленная
различным порядком связи атомов углерода.
Простейший
пример — бутан СН3—СН2—СН2—СН3 и изобутан (СН3)3СН.
Другие примеры: антрацен и фенантрен (формулы
I и II, соответственно), циклобутан иметилциклопропан (III
и IV).
Валентная
изомерия
Валентная —
особый вид структурной изомерии, при
которой изомеры можно перевести друг
в друга лишь за счёт перераспределения
связей. Например, валентными
изомерами бензола (V)
являются бицикло[2.2.0]гекса-2,5-диен (VI,
«бензол Дьюара»), призман (VII,
«бензол Ладенбурга»), бензвален (VIII).
Изомерия
функциональной группы (межклассовая
изомерия)[
Различается
характером функциональной группы;
например, этанол (CH3—CH2—OH)
и диметиловый
эфир (CH3—O—CH3).
Изомерия
положения[
Тип
структурной изомерии, характеризующийся
различием положения одинаковых
функциональных групп или кратных связей
при одинаковом углеродном скелете.
Пример: 2-хлорбутановая кислота и
4-хлорбутановая кислота.
Метамерия
Метамерия —
вид структурной
изомерии,
для которого характерно различное
распределение углеродных атомов между
несколькими углеводородными радикалами,
разделенными в молекуле гетероатомом.
Метамерия известна в рядах
алифатических простых
эфиров, сложных
эфиров, тиоспиртов и аминов.
В настоящее время термин используется
редко.
На
данный вид изомерии ещё указывал
А. М. Бутлеров, называя его «изомерия
нецельных структур».
Пример:
CH3CH2OCH2CH3 — диэтиловый
эфир и
CH3OCH2CH2CH3 —
метилпропиловый эфир
5. Пространственная изомерия
В
молекулах, содержащих
только одинарные связи, при
комнатной температуре
возможно почти свободное
вращение фрагментов молекулы
вокруг связей, и, например,
все изображения формул
1,2-дихлорэтана равноценны.
Рис. 4
Рис.
4. Положение атомов хлора
вокруг одинарной связи
Если
же вращение затруднено,
например, в циклической
молекуле или при двойной
связи, то возникает геометрическая
или цис-транс изомерия. В
цис-изомерах заместители
находятся по одну сторону
плоскости цикла или двойной
связи, в транс-изомерах – по разные
стороны.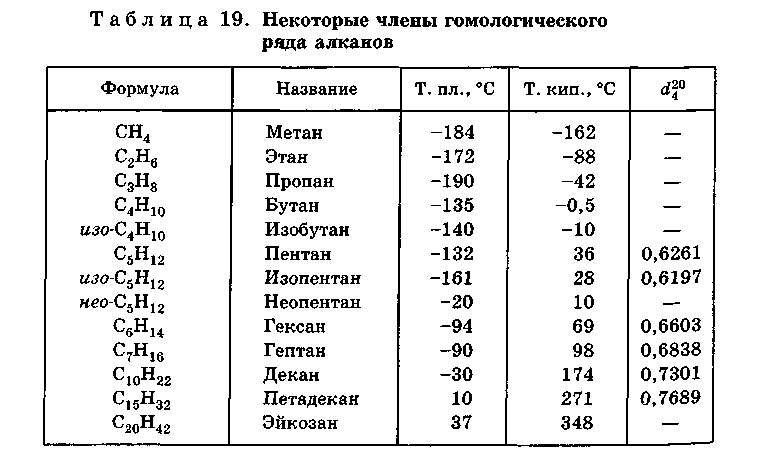
Цис-транс
изомеры существуют в том
случае, когда с атомом углерода
связаны два
разных заместителя.
Рис. 5.
Рис.
5. Цис- и транс- изомеры
Характерные химические свойства углеводородов. Механизмы реакций | ЕГЭ по химии
Характерные химические свойства углеводородов: алканов, алкенов, диенов, алкинов, ароматических углеводородов
Алканы
Алканы — углеводороды, в молекулах которых атомы связаны одинарными связями и которые соответствуют общей формуле $С_{n}Н_{2n+2}$.
Гомологический ряд метана
Как вы уже знаете, гомологи — это вещества, сходные по строению и свойствам и отличающиеся на одну или более групп $СН_2$.
Предельные углеводороды составляют гомологический ряд метана.
Изомерия и номенклатура
Для алканов характерна так называемая структурная изомерия. Структурные изомеры отличаются друг от друга строением углеродного скелета. Как вам уже известно, простейший алкан, для которого характерны структурные изомеры, — это бутан:
Как вам уже известно, простейший алкан, для которого характерны структурные изомеры, — это бутан:
Рассмотрим подробнее для алканов основы номенклатуры ИЮПАК:
1. Выбор главной цепи.
Формирование названия углеводорода начинается с определения главной цепи — самой длинной цепочки атомов углерода в молекуле, которая является как бы ее основой.
2. Нумерация атомов главной цепи.
Атомам главной цепи присваивают номера. Нумерация атомов главной цепи начинается с того конца, к которому ближе стоит заместитель (структуры А, Б). Если заместители находятся на равном удалении от конца цепи, то нумерация начинается от того конца, при котором их больше (структура В). Если различные заместители находятся на равном удалении от концов цепи, то нумерация начинается с того конца, к которому ближе старший (структура Г). Старшинство углеводородных заместителей определяется по тому, в каком порядке следует в алфавите буква, с которой начинается их название: метил (—$СН_3$), затем пропил ($—СН_2—СН_2—СН_3$), этил ($—СН_2—СН_3$) и т. д.
д.
Обратите внимание на то, что название заместителя формируется заменой суффикса -ан на суффикс -ил в названии соответствующего алкана.
3. Формирование названия.
В начале названия указывают цифры — номера атомов углерода, при которых находятся заместители. Если при данном атоме находятся несколько заместителей, то соответствующий номер в названии повторяется дважды через запятую ($2.2-$). После номера через дефис указывают количество заместителей (ди — два, три — три, тетра — четыре, пента — пять) и название заместителя (метил, этил, пропил). Затем без пробелов и дефисов — название главной цепи. Главная цепь называется как углеводород — член гомологического ряда метана (метан, этан, пропан и т. д.).
Названия веществ, структурные формулы которых приведены выше, следующие:
— структура А: $2$-метилпропан;
— структура Б: $3$-этилгексан;
— структура В: $2,2,4$-триметилпентан;
— структура Г: $2$-метил $4$-этилгексан.
Физические и химические свойства алканов
Физические свойства. Первые четыре представителя гомологического ряда метана — газы. Простейший из них — метан — газ без цвета, вкуса и запаха (запах газа, почувствовав который, надо звонить $104$, определяется запахом меркаптанов — серосодержащих соединений, специально добавляемых к метану, используемому в бытовых и промышленных газовых приборах, для того, чтобы люди, находящиеся рядом с ними, могли по запаху определить утечку).
Углеводороды состава от $С_5Н_{12}$ до $С_{15}Н_{32}$ — жидкости; более тяжелые углеводороды — твердые вещества.
Температуры кипения и плавления алканов постепенно увеличиваются с возрастанием длины углеродной цепи. Все углеводороды плохо растворяются в воде, жидкие углеводороды являются распространенными органическими растворителями.
Химические свойства.
1. Реакции замещения. Наиболее характерными для алканов являются реакции свободнорадикального замещения, в ходе которого атом водорода замещается на атом галогена или какую-либо группу.
Приведем уравнения наиболее характерных реакций.
Галогенирование:
$CH_4+Cl_2→CH_3Cl+HCl$.
В случае избытка галогена хлорирование может пойти дальше, вплоть до полного замещения всех атомов водорода на хлор:
$CH_3Cl+Cl_2→HCl+{CH_2Cl_2}↙{\text»дихлорметан(хлористый метилен)»}$,
$CH_2Cl_2+Cl_2→HCl+{CHСl_3}↙{\text»трихлорметан(хлороформ)»}$,
$CHCl_3+Cl_2→HCl+{CCl_4}↙{\text»тетрахлорметан(четыреххлористый углерод)»}$.
Полученные вещества широко используются как растворители и исходные вещества в органических синтезах.
2. Дегидрирование (отщепление водорода). В ходе пропускания алканов над катализатором ($Pt, Ni, Al_2O_3, Cr_2O_3$) при высокой температуре ($400–600°С$) происходит отщепление молекулы водорода и образование алкена:
$CH_3—CН_3→СH_2=CH_2+Н_2↑$
3. Реакции, сопровождающиеся разрушением углеродной цепи. Все предельные углеводороды горят с образованием углекислого газа и воды. Газообразные углеводороды, смешанные с воздухом в определенных соотношениях, могут взрываться. Горение предельных углеводородов — это свободнорадикальная экзотермическая реакция, которая имеет очень большое значение при использовании алканов в качестве топлива:
Горение предельных углеводородов — это свободнорадикальная экзотермическая реакция, которая имеет очень большое значение при использовании алканов в качестве топлива:
$СН_4+2О_2→СО_2+2Н_2O+880 кДж.$
В общем виде реакцию горения алканов можно записать следующим образом:
$C_{n}H_{2n+2}+({3n+1}/{2})O_2→nCO_2+(n+1)H_2O$
Термическое расщепление углеводородов:
$C_{n}H_{2n+2}{→}↖{400-500°C}C_{n-k}H_{2(n-k)+2}+C_{k}H_{2k}$
Процесс протекает по свободнорадикальному механизму. Повышение температуры приводит к гомолитическому разрыву углерод-углеродной связи и образованию свободных радикалов:
$R—CH_2CH_2:CH_2—R→R—CH_2CH_2·+·CH_2—R$.
Эти радикалы взаимодействуют между собой, обмениваясь атомом водорода, с образованием молекулы алкана и молекулы алкена:
$R—CH_2CH_2·+·CH_2—R→R—CH=CH_2+CH_3—R$.
Реакции термического расщепления лежат в основе промышленного процесса — крекинга углеводородов. Этот процесс является важнейшей стадией переработки нефти. 3$-гибридизации. Молекулы этих веществ построены при помощи ковалентных неполярных $С—С$ (углерод — углерод) связей и слабополярных $С—Н$ (углерод — водород) связей. В них нет участков с повышенной и с пониженной электронной плотностью, легко поляризуемых связей, т.е. таких связей, электронная плотность в которых может смещаться под действием внешних факторов (электростатических полей ионов). Следовательно, алканы не будут реагировать с заряженными частицами, т.к. связи в молекулах алканов не разрываются по гетеролитическому механизму.
3$-гибридизации. Молекулы этих веществ построены при помощи ковалентных неполярных $С—С$ (углерод — углерод) связей и слабополярных $С—Н$ (углерод — водород) связей. В них нет участков с повышенной и с пониженной электронной плотностью, легко поляризуемых связей, т.е. таких связей, электронная плотность в которых может смещаться под действием внешних факторов (электростатических полей ионов). Следовательно, алканы не будут реагировать с заряженными частицами, т.к. связи в молекулах алканов не разрываются по гетеролитическому механизму.
Алкены
К непредельным относят углеводороды, содержащие в молекулах кратные связи между атомами углерода. Непредельными являются алкены, алкадиены (полиены), алкины. Непредельным характером обладают также циклические углеводороды, содержащие двойную связь в цикле (циклоалкены), а также циклоалканы с небольшим числом атомов углерода в цикле (три или четыре атома). Свойство непредельности связано со способностью этих веществ вступать в реакции присоединения, прежде всего водорода, с образованием предельных, или насыщенных, углеводородов — алканов.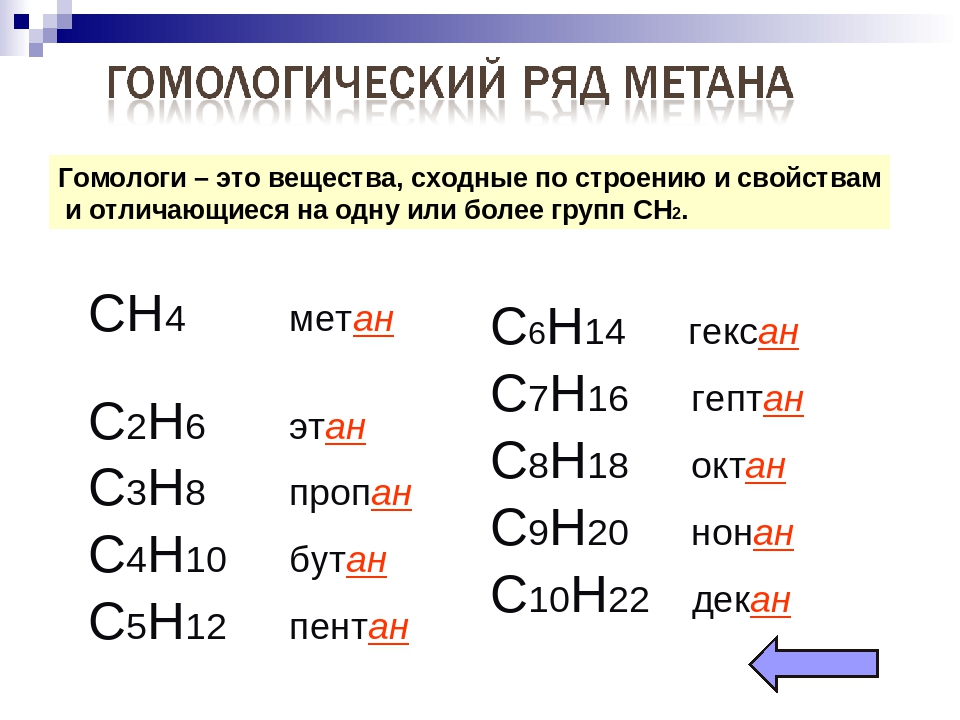
Алкены — ациклические углеводороды, содержащие в молекуле, помимо одинарных связей, одну двойную связь между атомами углерода и соответствующие общей формуле $С_{n}Н_{2n}$.
Свое второе название — олефины — алкены получили по аналогии с жирными непредельными кислотами (олеиновая, линолевая), остатки которых входят в состав жидких жиров — масел (от лат. oleum — масло).
Гомологический ряд этена
Неразветвленные алкены составляют гомологический ряд этена (этилена):
$С_2Н_4$ — этен, $С_3Н_6$ — пропен, $С_4Н_8$ — бутен, $С_5Н_{10}$ — пентен, $С_6Н_{12}$ — гексен и т. д.
Изомерия и номенклатура
Для алкенов, так же, как и для алканов, характерна структурная изомерия. Структурные изомеры отличаются друг от друга строением углеродного скелета. Простейший алкен, для которого характерны структурные изомеры, — это бутен:
Особым видом структурной изомерии является изомерия положения двойной связи:
$СН_3—{СН_2}↙{бутен-1}—СН=СН_2$ $СН_3—{СН=СН}↙{бутен-2}—СН_3$
Вокруг одинарной углерод-углеродной связи возможно практически свободное вращение атомов углерода, поэтому молекулы алканов могут приобретать самую разнообразную форму.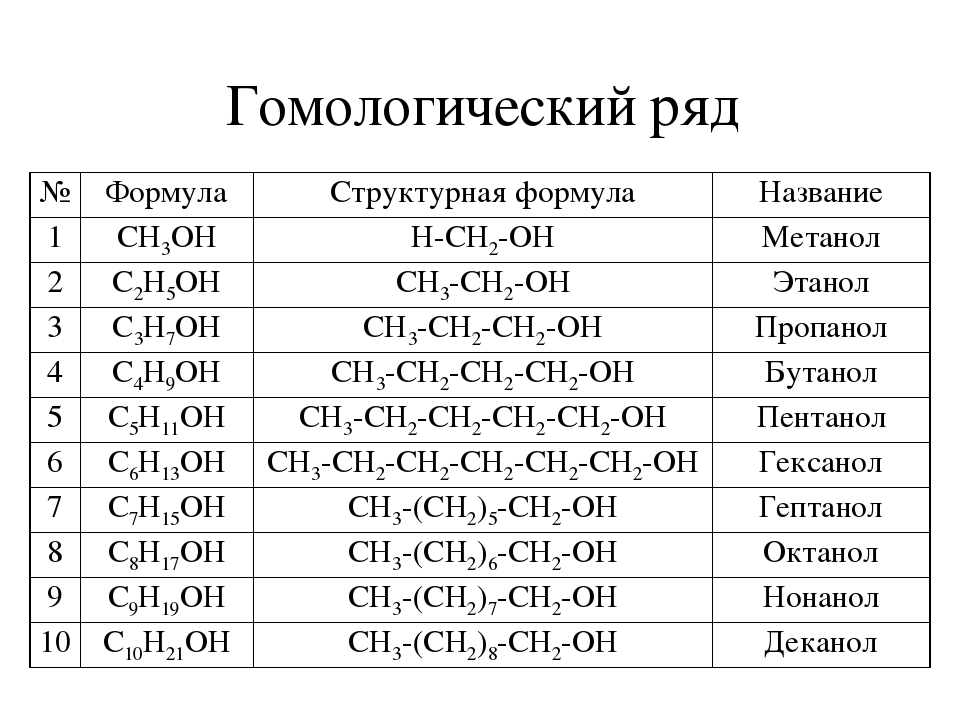 Вращение вокруг двойной связи невозможно, что приводит к появлению у алкенов еще одного вида изомерии — геометрической, или цис-транс изомерии.
Вращение вокруг двойной связи невозможно, что приводит к появлению у алкенов еще одного вида изомерии — геометрической, или цис-транс изомерии.
Цис-изомеры отличаются от транс-изомеров пространственным расположением фрагментов молекулы (в данном случае метильных групп) относительно плоскости $π$-связи, а следовательно, и свойствами.
Алкены изомерны циклоалканам (межклассовая изомерия), например:
Номенклатура алкенов, разработанная ИЮПАК, схожа с номенклатурой алканов.
1. Выбор главной цепи.
Образование названия углеводорода начинается с определения главной цепи — самой длинной цепочки атомов углерода в молекуле. В случае алкенов главная цепь должна содержать двойную связь.
2. Нумерация атомов главной цепи.
Нумерация атомов главной цепи начинается с того конца, к которому ближе находится двойная связь. Например, правильное название соединения:
$5$-метилгексен-$2$, а не $2$-метилгексен-$4$, как можно было бы предположить.
Если по положению двойной связи нельзя определить начало нумерации атомов в цепи, то его определяет положение заместителей, так же, как для предельных углеводородов.
3. Формирование названия.
Названия алкенов формируются так же, как и названия алканов. В конце названия указывают номер атома углерода, у которого начинается двойная связь, и суффикс, обозначающий принадлежность соединения к классу алкенов, — -ен.
Например:
Физические и химические свойства алкенов
Физические свойства. Первые три представителя гомологического ряда алкенов — газы; вещества состава $С_5Н_{10}$ – $С_{16}Н_{32}$ — жидкости; высшие алкены — твердые вещества.
Температуры кипения и плавления закономерно повышаются при увеличении молекулярной массы соединений.
Химические свойства.
Реакции присоединения. Напомним, что отличительной чертой представителей непредельных углеводородов — алкенов является способность вступать в реакции присоединения. Большинство этих реакций протекает по механизму электрофильного присоединения.
1. Гидрирование алкенов. Алкены способны присоединять водород в присутствии катализаторов гидрирования, металлов — платины, палладия, никеля:
$CH_3—CH_2—CH=CH_2+H_2{→}↖{Pt}CH_3—CH_2—CH_2—CH_3$.
Эта реакция протекает при атмосферном и повышенном давлении и не требует высокой температуры, т.к. является экзотермической. При повышении температуры на тех же катализаторах может пойти обратная реакция — дегидрирование.
2. Галогенирование (присоединение галогенов). Взаимодействие алкена с бромной водой или раствором брома в органическом растворителе ($CCl_4$) приводит к быстрому обесцвечиванию этих растворов в результате присоединения молекулы галогена к алкену и образования дигалоген алканов:
$СН_2=СН_2+Br_2→CH_2Br—CH_2Br$.
3. Гидрогалогенирование (присоединение галогеноводорода).
$CH_3-{CH}↙{пропен}=CH_2+HBr→CH_3-{CHBr}↙{2-бромпропен}-CH_3$
Эта реакция подчиняется правилу Марковникова:
При присоединении галогеноводорода к алкену водород присоединяется к более гидрированному атому углерода, т.е. атому, при котором находится больше атомов водорода, а галоген — к менее гидрированному.
4. Гидратация (присоединение воды).
Гидратация алкенов приводит к образованию спиртов. Например, присоединение воды к этену лежит в основе одного из промышленных способов получения этилового спирта:
${CH_2}↙{этен}=CH_2+H_2O{→}↖{t,H_3PO_4}CH_3-{CH_2OH}↙{этанол}$
Обратите внимание на то, что первичный спирт (с гидроксогруппой при первичном углероде) образуется только при гидратации этена. При гидратации пропена или других алкенов образуются вторичные спирты.
Эта реакция протекает также в соответствии с правилом Марковникова — катион водорода присоединяется к более гидрированному атому углерода, а гидроксогруппа — к менее гидрированному.
5. Полимеризация. Особым случаем присоединения является реакция полимеризации алкенов:
$nCH_2{=}↙{этен}CH_2{→}↖{УФ-свет,R}(…{-CH_2-CH_2-}↙{полиэтилен}…)_n$
Эта реакция присоединения протекает по свободнорадикальному механизму.
6. Реакция окисления.
Как и любые органические соединения, алкены горят в кислороде с образованием $СО_2$ и $Н_2О$:
$СН_2=СН_2+3О_2→2СО_2↑+2Н_2О$.
В общем виде:
$C_{n}H_{2n}+{3n}/{2}O_2→nCO_2↑+nH_2O$
В отличие от алканов, которые устойчивы к окислению в растворах, алкены легко окисляются под действием растворов перманганата калия. В нейтральных или щелочных растворах происходит окисление алкенов до диолов (двухатомных спиртов), причем гидроксильные группы присоединяются к тем атомам, между которыми до окисления существовала двойная связь:
Алкадиены (диеновые углеводороды)
Алкадиены — ациклические углеводороды, содержащие в молекуле, помимо одинарных связей, две двойные связи между атомами углерода и соответствующие общей формуле $С_{n}Н_{2n-2}$.
В зависимости от взаимного расположения двойных связей различают три вида диенов:
— алкадиены с кумулированным расположением двойных связей:
$CH_2=C=CH_2$;
— алкадиены с сопряженными двойными связями;
$CH_2=CH—CH=CH_2$;
— алкадиены с изолированными двойными связями
$CH_2=CH—CH_2—CH=CH_2$.
Эти все три вида алкадиенов существенно отличаются друг от друга по строению и свойствам. Центральный атом углерода (атом, образующий две двойные связи) в алкадиенах с кумулированными связями находится в состоянии $sp$-гибридизации. Он образует две $σ$-связи, лежащие на одной прямой и направленные в противоположные стороны, и две $π$-связи, лежащие в перпендикулярных плоскостях. $π$-Связи образуются за счет негибридизированных р-орбиталей каждого атома углерода. Свойства алкадиенов с изолированными двойными связями весьма специфичны, т.к. сопряженные $π$-связи существенно влияют друг на друга.
р-Орбитали, образующие сопряженные $π$-связи, составляют практически единую систему (ее называют $π$-системой), т.к. р-орбитали соседних $π$-связей частично перекрываются.
Изомерия и номенклатура
Для алкадиенов характерна как структурная изомерия, так и цис-, транс-изомерия.
Структурная изомерия.
— изомерия углеродного скелета:
— изомерия положения кратных связей:
${CH_2=CH—CH=CH_2}↙{бутадиен-1,3}$ ${CH_2=C=CH—CH_3}↙{бутадиен-1,2}$
Цис-, транс-изомерия (пространственная и геометрическая)
Например:
Алкадиены изомерны соединениям классов алкинов и циклоалкенов.
При формировании названия алкадиена указывают номера двойных связей. Главная цепь должна обязательно содержать две кратные связи.
Например:
Физические и химические свойства алкадиенов
Физические свойства.
В обычных условиях пропандиен-1,2, бутадиен-1,3 — газы, 2-метилбутадиен-1,3 — летучая жидкость. Алкадиены с изолированными двойными связями (простейший из них — пентадиен-1,4) — жидкости. Высшие диены — твердые вещества.
Химические свойства.
Химические свойства алкадиенов с изолированными двойными связями мало отличаются от свойств алкенов. Алкадиены с сопряженными связями обладают некоторыми особенностями.
1. Реакции присоединения. Алкадиены способны присоединять водород, галогены, галогеноводороды.
Особенностью присоединения к алкадиенам с сопряженными связями является способность присоединять молекулы как в положениях 1 и 2, так и в положениях 1 и 4.
Соотношение продуктов зависит от условий и способа проведения соответствующих реакций.
2. Реакция полимеризации. Важнейшим свойством диенов является способность полимеризоваться под воздействием катионов или свободных радикалов. Полимеризация этих соединений является основой синтетических каучуков:
$nCH_2={CH—CH=CH_2}↙{бутадиен-1,3}→{(… —CH_2—CH=CH—CH_2— …)_n}↙{\text»синтетический бутадиеновый каучук»}$.
Полимеризация сопряженных диенов протекает как 1,4-присоединение.
В этом случае двойная связь оказывается центральной в звене, а элементарное звено, в свою очередь, может принимать как цис-, так и транс-конфигурацию.
Алкины
Алкины — ациклические углеводороды, содержащие в молекуле, помимо одинарных связей, одну тройную связь между атомами углерода и соответствующие общей формуле $С_{n}Н_{2n-2}$.
Гомологический ряд этина
Неразветвленные алкины составляют гомологический ряд этина (ацетилена):
$С_2Н_2$ — этин, $С_3Н_4$ — пропин, $С_4Н_6$ — бутин, $С_5Н_8$ — пентин, $С_6Н_{10}$ — гексин и т. д.
Изомерия и номенклатура
Для алкинов, так же как и для алкенов, характерна структурная изомерия: изомерия углеродного скелета и изомерия положения кратной связи. Простейший алкин, для которого характерны структурные изомеры положения кратной связи класса алкинов, — это бутин:
$СН_3—{СН_2}↙{бутин-1}—С≡СН$ $СН_3—{С≡С}↙{бутин-2}—СН_3$
Изомерия углеродного скелета у алкинов возможна, начиная с пентина:
Так как тройная связь предполагает линейное строение углеродной цепи, геометрическая (цис-, транс-) изомерия для алкинов невозможна.
Наличие тройной связи в молекулах углеводородов этого класса отражается суффиксом -ин, а ее положение в цепи — номером атома углерода.
Например:
Алкинам изомерны соединения некоторых других классов. Так, химическую формулу $С_6Н_{10}$ имеют гексин (алкин), гексадиен (алкадиен) и циклогексен (циклоалкен):
Физические и химические свойства алкинов
Физические свойства. Температуры кипения и плавления алкинов, так же, как и алкенов, закономерно повышаются при увеличении молекулярной массы соединений.
Алкины имеют специфический запах. Они лучше растворяются в воде, чем алканы и алкены.
Химические свойства.
Реакции присоединения. Алкины относятся к непредельным соединениям и вступают в реакции присоединения. В основном это реакции электрофильного присоединения.
1. Галогенирование (присоединение молекулы галогена). Алкин способен присоединить две молекулы галогена (хлора, брома):
$CH≡CH+Br_2→{CHBr=CHBr}↙{1,2-дибромэтан},$
$CHBr=CHBr+Br_2→{CHBr_2-CHBr_2}↙{1,1,2,2-тетрабромэтан}$
2. Гидрогалогенирование (присоединение галогеноводорода). Реакция присоединения галогеноводорода, протекающая по электрофильному механизму, также идет в две стадии, причем на обеих стадиях выполняется правило Марковникова:
$CH_3-C≡CH+Br→{CH_3-CBr=CH_2}↙{2-бромпропен},$
$CH_3-CBr=CH_2+HBr→{CH_3-CHBr_2-CH_3}↙{2,2-дибромпропан}$
3. Гидратация (присоединение воды). Боль шое значение для промышленного синтеза кетонов и альдегидов имеет реакция присоединения воды (гидратация), которую называют реакцией Кучерова:
4. Гидрирование алкинов. Алкины присоединяют водород в присутствии металлических катализаторов ($Pt, Pd, Ni$):
$R-C≡C-R+H_2{→}↖{Pt}R-CH=CH-R,$
$R-CH=CH-R+H_2{→}↖{Pt}R-CH_2-CH_2-R$
Так как тройная связь содержит две реакционноспособные $π$-связи, алканы присоединяют водород ступенчато:
1) тримеризация.
При пропускании этина над активированным углем образуется смесь продуктов, одним из которых является бензол:
2) димеризация.
Помимо тримеризации ацетилена, возможна его димеризация. Под действием солей одновалентной меди образуется винилацетилен:
$2HC≡CH→{HC≡C-CH=CH_2}↙{\text»бутен-1-ин-3(винилацетилен)»}$
Это вещество используется для получения хлоропрена:
$HC≡C-CH=CH_2+HCl{→}↖{CaCl}H_2C={CCl-CH}↙{хлоропрен}=CH_2$
полимеризацией которого получают хлоропреновый каучук:
$nH_2C=CCl-CH=CH_2→(…-H_2C-CCl=CH-CH_2-…)_n$
Окисление алкинов.
Этин (ацетилен) горит в кислороде с выделением очень большого количества теплоты:
$2C_2H_2+5O_2→4CO_2↑+2H_2O+2600кДж$ На этой реакции основано действие кислородно-ацетиленовой горелки, пламя которой имеет очень высокую температуру (более $3000°С$), что позволяет использовать ее для резки и сварки металлов.
На воздухе ацетилен горит коптящим пламенем, т.к. содержание углерода в его молекуле выше, чем в молекулах этана и этена.
Алкины, как и алкены, обесцвечивают подкисленные растворы перманганата калия; при этом происходит разрушение кратной связи.
Ионный (правило В. В. Марковникова) и радикальный механизмы реакции в органической химии
Типы химических реакций в органической химии
Реакции органических веществ можно формально разделить на четыре основных типа: замещения, присоединения, отщепления (элиминирования) и перегруппировки (изомеризации). Очевидно, что все многообразие реакций органических соединений невозможно свести к предложенной классификации (например, реакции горения). Однако такая классификация поможет установить аналогии с уже знакомыми вам из курса неорганической химии реакциями, протекающими между неорганическими веществами.
Как правило, основное органическое соединение, участвующее в реакции, называют субстратом, а другой компонент реакции условно рассматривают как реагент.
Реакции замещения
Реакции, в результате которых осуществляется замена одного атома или группы атомов в исходной молекуле (субстрате) на другие атомы или группы атомов, называются реакциями замещения.
В реакции замещения вступают предельные и ароматические соединения, такие как алканы, циклоалканы или арены.
Приведем примеры таких реакций.
Под действием света атомы водорода в молекуле метана способны замещаться на атомы галогена, например, на атомы хлора:
$CH_4+Cl_2→CH_3Cl+HCl$
Другим примером замещения водорода на галоген является превращение бензола в бромбензол:
Уравнение этой реакции может быть записано иначе:
При этой форме записи реагенты, катализатор, условия проведения реакции записывают над стрелкой, а неорганические продукты реакции — под ней.
Реакции присоединения
Реакции, в результате которых две или более молекул реагирующих веществ соединяются в одну, называют реакциями присоединения.
В реакции присоединения вступают ненасыщенные соединения, такие как алкены или алкины.
В зависимости от того, какая молекула выступает в качестве реагента, различают гидрирование (или восстановление), галогенирование, гидрогалогенирование, гидратацию и другие реакции присоединения. Каждая из них требует определенных условий.
1. Гидрирование — реакция присоединения молекулы водорода по кратной связи:
$CH_3{-CH=}↙{\text»пропен»}CH_2+H_2{→}↖{Pt}CH_3{-CH_2-}↙{\text»пропан»}-CH_3$
2. Гидрогалогенирование — реакция присоединения галогеноводорода (гидрохлорирование):
${CH_2=}↙{\text»этен»}CH_2+HCl→CH_3{-CH_2-}↙{\text»хлорэтан»}-Cl$
3. Галогенирование — реакция присоединения галогена:
${CH_2=}↙{\text»этен»}CH_2+Cl_2→{CH_2Cl-CH_2Cl}↙{\text»1.2-дихлорэтан»}$
4. Полимеризация — особый тип реакций присоединения, в ходе которых молекулы вещества с небольшой молекулярной массой соединяются друг с другом с образованием молекул вещества с очень высокой молекулярной массой — макромолекул.
Реакции полимеризации — это процессы соединения множества молекул низкомолекулярного вещества (мономера) в крупные молекулы (макромолекулы) полимера.
Примером реакции полимеризации может служить получение полиэтилена из этилена (этена) под действием ультрафиолетового излучения и радикального инициатора полимеризации $R:$
${nCH_2=}↙{\text»этен»}CH_2{→}↖{\text»УФ-свет,R»}{(…-CH_2-CH_2-…)_n}↙{\text»полиэтилен»}$
Наиболее характерная для органических соединений ковалентная связь образуется при перекрывании атомных орбиталей и образовании общих электронных пар. В результате этого образуется общая для двух атомов орбиталь, на которой находится общая электронная пара. При разрыве связи судьба этих общих электронов может быть разной.
Типы реакционноспособных частиц в органической химии
Орбиталь с неспаренным электроном, принадлежащая одному атому, может перекрываться с орбиталью другого атома, на которой также находится неспаренный электрон. При этом происходит образование ковалентной связи по обменному механизму:
$H•+•H→H:H,$ или $H-H$
Обменный механизм образования ковалентной связи реализуется в том случае, если общая электронная пара образуется из неспаренных электронов, принадлежащих разным атомам.
Процессом, противоположным образованию ковалентной связи по обменному механизму, является разрыв связи, при котором к каждому атому отходит по одному электрону. В результате этого образуются две незаряженные частицы, имеющие неспаренные электроны:
$R:R=R•+R•$
Такие частицы называются свободными радикалами.
Свободные радикалы — атомы или группы атомов, имеющие неспаренные электроны.
Реакции, которые протекают под действием и при участии свободных радикалов, называются свободнорадикальными реакциями.{-}$ (ацетат-анион).
Частицы, имеющие незаполненную орбиталь, напротив, будут стремиться заполнить ее и, следовательно, будут притягиваться к участкам молекул, на которых присутствует повышенная электронная плотность, отрицательный заряд, неподеленная электронная пара. Они являются электрофилами, «друзьями» электрона, отрицательного заряда или частиц с повышенной электронной плотностью.
Электрофилы — катионы или молекулы, имеющие незаполненную эле к трон ную орбиталь, стремящиеся к заполнению ее электронами, так как это приводит к более выгодной электронной конфигурации атома.
Примеры электрофилов: $NO_2$ (нитрогруппа), —$СООН$ (карбоксил), —$СN$ (нитрильная группа), —$СОН$ (альдегидная группа).
Не любая частица с незаполненной орбиталью является электрофилом. Так, например, катионы щелочных металлов имеют конфигурацию инертных газов и не стремятся к приобретению электронов, так как имеют низкое сродство к электрону. Из этого можно сделать вывод, что, несмотря на наличие у них незаполненной орбитали, подобные частицы не будут являться электрофилами.
Основные механизмы протекания реакций
Мы выделили три основных типа реагирующих частиц — свободные радикалы, электрофилы, нуклеофилы — и три соответствующих им типа механизма реакций:
— свободнорадикальные;
— электрофильные;
— нуклеофильные.
Кроме классификации реакций по типу реагирующих частиц, в органической химии различают четыре вида реакций по принципу изменения состава молекул: присоединения, замещения, отщепления, или элиминирования (от лат. eliminaue — удалять, отщеплять) и перегруппировки. Так как присоединение и замещение могут происходить под действием всех трех типов реакционноспособных частиц, можно выделить несколько основных механизмов протекания реакций.
1. Свободнорадикальное замещение:
${CH_4}↙{\text»метан»}+Br_2{→}↖{\text»УФ-свет»}{CH_3Br}↙{\text»бромметан»}+HBr$
2. Свободнорадикальное присоединение:
$nCH_2=CH_2{→}↖{\text»УФ-свет,R»}(…-CH_2-CH_2-…)_n$
3. Электрофильное замещение:
4. Электрофильное присоединение:
$CH_3-{CH=}↙{\text»пропен»}CH_2+HBr{→}↖{\text»раствор»}{CH_3-CHBr-CH_3}↙{\text»2-бромпропан»}$
$СH_3{-C≡}↙{\text»пропин»}CH+Cl_2{→}↖{\text»раствор»}{CH_3-CCl=CHCl}↙{\text»1,2-дихлорпропен»}$
5. Нуклеофильное присоединение:
Кроме того, мы рассмотрим реакции отщепления, или элиминирования, которые идут под воздействием нуклеофильных частиц — оснований.
6. Элиминирование:
$СH_3-CHBr-CH_3+NaOH{→}↖{\text»спиртовой раствор»}CH_3-CH=CH_2+NaBr+H_2O$
Можно считать, что в этой реакции происходит отщепление молекулы бромводорода от молекулы 2-бромпропана. В присутствии щелочи образуются бромид натрия и вода.
Правило В. В. Марковникова
Отличительной чертой алкенов (непредельных углеводородов) является способность вступать в реакции присоединения. Большинство этих реакций протекает по механизму электрофильного присоединения.
Гидрогалогенирование (присоединение галогеноводо рода):
$СH_3{-CH-}↙{\text»пропен»}CH_2+HBr→CH_3{-CHBr-CH_3}↙{\text»2-бромпропан»}$
Эта реакция подчиняется правилу В. В. Марковникова: при присоединении галогенводорода к алкену водород присоединяется к более гидрированному атому углерода, т.е. атому, при котором находится больше атомов водорода, а галоген — к менее гидрированному.
Гомология (биология) — Энциклопедия Нового Света
Кости человеческого запястья (запястья), ладони (пястные кости), а также пальцев и большого пальца (пальцы) имеют сильное сходство с очень разнообразными гомологичными структурами в передних конечностях и » руки »млекопитающих.
В биологии гомология обычно определяется как любое сходство между структурами организмов в разных таксонах, которое происходит из сходных структур их общего предка. Примеры гомологии встречаются на разных уровнях организации.Целые анатомические структуры, сходные в разных биологических таксонах (видах, родах и т. Д.), Будут называться гомологичными, если их общий предок также демонстрирует аналогичную анатомическую структуру. Тесно коррелированные частичные последовательности в ДНК или белке могут быть помечены аналогичным образом, если общее происхождение считается причиной.
Такое определение гомологии является переопределением классического понимания этого термина, которое предшествовало теории эволюции Чарльза Дарвина, придуманной Ричардом Оуэном в 1840-х годах.Исторически гомология определялась как сходство в структуре и положении, например, рисунок костей в крыле летучей мыши и в плавнике морской свиньи (Wells, 2000). Исторически и до сих пор термин аналогия обозначал функциональное сходство, например, крылья птицы и бабочки.
Дарвин утверждал, что гомология в классическом смысле является доказательством эволюции путем общего происхождения (теория происхождения с модификацией), предполагая, что новые формы развиваются на основе более ранних стадий.Его доказательства включали аналогичные скелетные структуры (использующие одни и те же кости) передних конечностей людей, летучих мышей, птиц, кошек и морских свиней.
Однако открытие явных исключений из предпосылки Дарвина означало, что структурное сходство больше не могло использоваться в качестве доказательства общего предка. Одним из таких исключений является поразительное структурное сходство между глазами человека и осьминога, двух организмов, чей дальний общий предок, как полагают, не имел таких глаз (Ogura, Ikeo, and Gojobori, 2004).Если общее происхождение должно быть установлено отдельно от структурного сходства, прежде чем структуры можно будет назвать гомологичными, то использование сходства двух структур в качестве доказательства их эволюционного происхождения, включая их общее происхождение, повлечет за собой использование круговых рассуждений.
Слово , гомологичное , происходит от древнегреческого ομολογειν, «соглашаться». Термин гомология также используется в неэволюционном смысле в отношении гомологичных хромосом, означая пару неидентичных хромосом из диплоидного организма, которые могут образовывать пары (синапсы) во время мейоза, или участки хромосом с одним и тем же набором генов.
Гомология структур в эволюции
Гомология в классическом смысле, как сходство в структуре и положении анатомических особенностей между разными организмами, была важным доказательством, используемым Дарвином. Однако теперь он был переопределен как структуры, восходящие к общим предкам. Майр (1982) утверждает:
После 1859 г. существовало только одно определение гомологичного, имеющее биологический смысл… Атрибуты двух организмов являются гомологичными, если они происходят из эквивалентных характеристик общего предка.
Гомология отличается от аналогии. Например, крылья насекомых, крылья летучих мышей и крылья птиц аналогичны, но не гомологичны; это явление называется «гомоплазия». Эти похожие структуры эволюционировали по разным путям развития в процессе, названном конвергентной эволюцией.
Уэллс (2000) отмечает, что гомология, как она теперь определена, больше не может использоваться в качестве доказательства эволюции из-за круговой аргументации. Более того, он находит, что даже сохранение додарвиновского определения гомологии как структурного сходства проблематично из-за таких факторов, как несоответствие между гомологией и путями развития, поскольку есть примеры подобных структур во взрослых формах, где предполагается, что предки действительно являются продуктами. кардинально разных процессов развития.Точно так же он отмечает, что некоторые предполагаемые гомологичные структуры восходят к разным генам.
Гомология: пентадактильная конечность
Принцип гомологии, проиллюстрированный адаптивным излучением передней конечности млекопитающих. Все они соответствуют основному паттерну пентадактиля, но модифицированы для разных целей. Третья пястная кость на всем протяжении заштрихована; плечо заштриховано.
Структура костей конечностей, известная как пентадактильная конечность, является примером гомологичных структур. Этот паттерн встречается у всех классов четвероногих ( и.е. от земноводных до млекопитающих). Его даже можно проследить до плавников некоторых ископаемых рыб, от которых, как считается, произошли первые земноводные. На конечности есть одна проксимальная кость (плечевая кость), две дистальные кости (лучевая и локтевая), ряд запястных костей (кости запястья), за которыми следуют пять рядов пястных костей (кости ладони) и фаланги (пальцы). У четвероногих фундаментальные структуры конечностей пентадактилей одинаковы, что указывает на то, что они произошли от общего предка. Но считается, что в ходе эволюции эти фундаментальные структуры претерпели изменения.Они стали внешне разными и не связанными друг с другом структурами, чтобы выполнять разные функции при адаптации к разным условиям и образу жизни. Это явление ярко проявляется на передних конечностях млекопитающих. Например:
- У обезьяны передние конечности сильно удлинены, чтобы образовывать хватательную руку для лазания и раскачивания между деревьями.
- У свиньи первая цифра потеряна, а вторая и пятая цифры уменьшены. Остальные два пальца длиннее и толще остальных и имеют копыто для поддержки тела.
- У лошади передние конечности приспособлены для поддержки и движения за счет большого удлинения третьего пальца, несущего копыто.
- У крота есть пара коротких лопатообразных передних конечностей для рытья.
- Муравьед использует свой увеличенный третий палец для сноса муравейников и гнезд термитов.
- У кита передние конечности становятся ластами для управления и поддержания равновесия во время плавания.
- У летучей мыши передние конечности превратились в крылья для полета за счет большого удлинения в четыре пальца, а крючковидный первый палец остается свободным, чтобы висеть на деревьях.
Гомология: ротовой аппарат насекомых
У насекомых основные строения ротового аппарата одинаковы, включая верхнюю губу (верхнюю губу), пару нижних челюстей, гипофаринкс (дно рта), пару верхних челюстей и верхнюю челюсть. губа. Эти структуры увеличиваются и видоизменяются; другие уменьшаются и теряются. Модификации позволяют насекомым использовать различные пищевые материалы:
(A) Примитивное состояние — кусание и жевание: например, кузнечик. Сильные нижние и верхние челюсти для манипуляций с пищей.
(B) Тикинг и укус: например, пчела. Губа долго поглощать нектар; челюсти жуют пыльцу и плесневой воск.
(C) Сосание: например, бабочка. Нижняя губа снижена; челюсти потеряны; maxillae длинные, образующие сосательную трубку.
(D) Пирсинг и сосание: например, самка комара. Верхняя и нижняя губа образуют трубку; нижние челюсти образуют пронзительные стилеты; верхняя губа рифленая для удержания других частей.
Гомология: придатки других членистоногих
Ротовые органы и усики насекомых считаются гомологами ног насекомых.Параллельное развитие наблюдается у некоторых паукообразных: передняя пара ног может быть видоизменена как аналоги антенн, особенно у хлыстовых скорпионов, которые ходят на шести ногах. Эти разработки подтверждают теорию о том, что сложные модификации часто возникают в результате дублирования компонентов, причем дубликаты изменяются в разных направлениях.
Гомология последовательностей в генетике
В генетике гомология измеряется путем сравнения последовательностей белков или ДНК, и гомологичные гены имеют высокую идентичность или сходство последовательностей, что подтверждает гипотезу о том, что они имеют общего предка.Гомология последовательностей также может указывать на общую функцию. Гомологичные хромосомы — это неидентичные хромосомы, которые могут образовывать пары (синапсы) во время мейоза и, как считается, имеют общее происхождение.
Гомология белков и ДНК часто делается на основании сходства последовательностей, особенно в биоинформатике. Например, в целом, если два гена имеют почти идентичную последовательность ДНК, вполне вероятно, что они гомологичны. Но сходство последовательностей может происходить от разных предков: короткие последовательности могут быть похожими случайно, и последовательности могут быть похожими, потому что оба были выбраны для связывания с конкретным белком, таким как фактор транскрипции.Такие последовательности похожи, но не гомологичны. Гомологичные участки последовательности также называют консервативными .
Иногда используется фраза «процент гомологии», но она неверна. «Процент идентичности» или «процент сходства» следует использовать для количественной оценки сходства между последовательностями биомолекул. Для двух встречающихся в природе последовательностей процент идентичности является фактическим измерением, тогда как гомология — это гипотеза, подтвержденная доказательствами. Однако можно сослаться на частичную гомологию, когда часть сравниваемых последовательностей (предполагается, что они) имеют общее происхождение, а остальные — нет.
Существует множество алгоритмов для кластеризации последовательностей белков в семейства последовательностей, которые представляют собой наборы взаимно гомологичных последовательностей.
Существует два типа гомологии последовательностей: ортологичная и паралогичная.
Ортология
Ортологи, или ортологичные гены — это любые гены разных видов, которые похожи друг на друга и, как считается, произошли от общего предка, независимо от их функций. Таким образом, ортологи разделены событием эволюционного видообразования: если ген существует у вида, и этот вид разделяется на два вида, то расходящиеся копии этого гена в результирующем виде являются ортологами.Термин «ортолог» был придуман в 1970 году.
Второе определение ортолога возникло для описания любых генов с очень похожими функциями у разных видов. Это отличается от исходного определения тем, что в нем нет утверждения об эволюционной связи или сходстве в последовательности или структуре.
Ортологические последовательности предоставляют полезную информацию при исследованиях таксономической классификации организмов. Паттерн генетической дивергенции можно использовать для отслеживания родства организмов.Два очень близких организма, вероятно, будут демонстрировать очень похожие последовательности ДНК между двумя ортологами. Напротив, организм, который далее эволюционно отделен от другого организма, вероятно, будет демонстрировать большее расхождение в последовательности изучаемых ортологов.
Паралогия
Гомологичные последовательности паралоги , если они были разделены событием дупликации гена; если ген в организме дублируется, чтобы занимать две разные позиции в одном геноме, то две копии являются паралогичными.
Набор последовательностей, которые являются паралогическими, называются паралогами друг друга. Паралоги обычно выполняют ту же или похожую функцию, но иногда их нет. Считается, что из-за отсутствия изначального селективного давления на одну копию дублированного гена эта копия может мутировать и приобретать новые функции.
Паралогичные последовательности дают полезное представление о том, как эволюционируют геномы. Гены, кодирующие миоглобин и гемоглобин, считаются древними паралогами.Точно так же четыре известных класса гемоглобинов (гемоглобин A, гемоглобин A2, гемоглобин S и гемоглобин F) считаются паралогами друг друга. Хотя каждый из этих генов выполняет одну и ту же основную функцию переноса кислорода, они немного различаются по функциям: гемоглобин плода (гемоглобин F) имеет более высокое сродство к кислороду, чем гемоглобин взрослого человека.
Другой пример можно найти у грызунов, таких как крысы и мыши. У грызунов есть пара генов инсулина, которые считаются паралогичными, хотя неясно, произошло ли какое-либо расхождение в функциях.
Паралогичные гены часто принадлежат к одному виду, но это не обязательно. Например, ген гемоглобина человека и ген миоглобина шимпанзе считаются паралогами. Это обычная проблема в биоинформатике; когда геномы разных видов были секвенированы и были обнаружены гомологичные гены, нельзя сразу сделать вывод, что эти гены имеют одинаковую или похожую функцию, поскольку они могут быть паралогами, функции которых разошлись.
Ксенология
Гомологи, полученные в результате горизонтального переноса генов между двумя организмами, называются ксенологами.Ксенологи могут иметь разные функции, если новая среда сильно отличается для горизонтально движущегося гена. В целом, однако, ксенологи обычно выполняют сходные функции у обоих организмов (NCBI, 2004).
Наборы гомологичных хромосом
Гомологичные хромосомы определяются как неидентичные хромосомы, которые могут образовывать пары (синапсы) во время мейоза (King and Stansfield, 1997). За исключением половых хромосом, гомологичные хромосомы имеют значительное сходство последовательностей по всей своей длине, обычно содержат одну и ту же последовательность генов и объединяются в пары, чтобы обеспечить правильное разъединение во время мейоза.На этом этапе хромосомы также могут подвергаться кроссинговеру. Могут быть некоторые вариации между генами по гомологам, приводящие к альтернативным формам или аллелям. Половые хромосомы имеют более короткую область сходства последовательностей. Основываясь на сходстве последовательностей и знаниях человека о биологии, считается, что они паралогичны.
Ссылки
- ДеПинна, М.С.С. 1991. Концепции и тесты гомологии в кладистической парадигме. Кладистика 7: 367-394.
- Дьюи, К., и Л. Пахтер. 2006. Эволюция на уровне нуклеотидов: проблема множественного выравнивания всего генома. Human Molecular Genetics 15 (Обзор, выпуск 1): R51-R56. Проверено 26 апреля 2007 г.
- Fitch, W. M. 2000. Гомология: личный взгляд на некоторые проблемы. Trends Genet. 16 (5): 227-31. Проверено 26 апреля 2007 года.
- Gegenbaur, G. 1898. Vergleichende Anatomie der Wirbelthiere. Лейпциг.
- Геккель, Е. 1866. Generelle Morphologie der Organismen. Bd 1-2. Берлин.
- King, R.C. и W. D. Stansfield. 1997. Словарь по генетике, 5-е изд. Нью-Йорк: Издательство Оксфордского университета. ISBN 0195094417.
- Национальный центр биотехнологической информации (NCBI). 2004. Только факты: базовое введение в науку, лежащую в основе ресурсов NCBI: систематика и молекулярная филогенетика. NCBI: A Science Primer . Проверено 26 апреля 2007 г.
- Mayr, E. 1982. Рост биологической мысли. Кембридж, Массачусетс: Издательство Гарвардского университета. ISBN 0674364457.
- Огура, А., Икео, К., Годжобори, Т. 2004. Сравнительный анализ экспрессии генов конвергентной эволюции глаза камеры у осьминога и человека. Genome Res. 14: 1555-1561. Проверено 28 января 2009 года.
- Оуэн, Р. 1847. Об архетипе и гомологиях скелета позвоночных. Лондон.
- Уэллс, Дж. 2000. Иконы эволюции . Вашингтон, округ Колумбия: Regnery Publishing.ISBN 0895262762.
Источники
Энциклопедия Нового Света писателей и редакторов переписали и завершили статью Википедия
в соответствии со стандартами New World Encyclopedia . Эта статья соответствует условиям лицензии Creative Commons CC-by-sa 3.0 (CC-by-sa), которая может использоваться и распространяться с указанием авторства. Кредит предоставляется в соответствии с условиями этой лицензии, которая может ссылаться как на участников New World Encyclopedia , так и на самоотверженных добровольцев Фонда Викимедиа.Чтобы процитировать эту статью, щелкните здесь, чтобы просмотреть список допустимых форматов цитирования. История более ранних вкладов википедистов доступна исследователям здесь:
История этой статьи с момента ее импорта в Энциклопедию Нового Света :
Примечание. могут применяться ограничения на использование отдельных изображений, на которые распространяется отдельная лицензия.
Гомология в развитии и развитие концепции гомологии1 | Интегративная и сравнительная биология
Абстрактные
Гомология — центральная концепция эволюции развития.Здесь я утверждаю, что гомологию следует объяснять в рамках исходных процессов развития и эволюции; развития, потому что это непосредственная причина морфологических признаков и эволюции, потому что она имеет дело с органическими преобразованиями и стабильностью. Это было уже признано Гансом Спеманом в 1915 году. В основополагающем эссе «История и критика концепции гомологии» Спеманн проанализировал историю и существующие проблемы концепции гомологии. Здесь я продолжу проект Спеманна и проанализирую некоторые из вкладов ХХ века в гомологию.Я закончу несколькими размышлениями о связях между процессами развития и гомологией и сделаю вывод, что процессы развития являются неотъемлемой частью (i) оценки гомологии, (ii) объяснения гомологии, (iii) происхождения эволюционных инноваций (зарождающиеся гомологи). ) и (iv) сами могут считаться гомологичными.
ВВЕДЕНИЕ
Семьдесят пять лет назад Ганс Спеманн завершил свое эссе Zur Geschichte und Kritik des Begriffs der Homologie (История и критика концепции гомологии) следующими, задним числом довольно пророческими словами.«Мы больше не верим, что можем сначала установить филогенетические отношения между животными, чтобы впоследствии вывести законы развития. Скорее мы начинаем понимать, что сначала мы должны определить эти законы, прежде чем мы сможем понять или даже установить морфологический ряд, который мы используем для классификации организмов (Spemann, 1915, стр. 84) ». В этом отрывке Спеманн доказывает центральную роль процессов развития для любого понимания морфологических трансформаций, , то есть ., Фенотипической эволюции, которые мы сейчас преследуем под заголовком Developmental Evolution или Evolution and Development. (для обозначения названий двух новых журналов, обслуживающих эту область, но также см. (Hall, 1992; Gilbert et al ., 1996; Raff, 1996; Gerhart and Kirschner, 1997).) Подчеркивая роль Даже для систематики, Спеманн выступал против широко распространенной тогда моды получения филогении на основе сравнительных анатомических и эмбриологических данных, которые в значительной степени опирались на то, что он определил как историческую концепцию гомологии (филогенетическая и онтогенетическая преемственность). В его эссе биологи-эволюционисты и эволюционисты продолжали интересоваться взаимосвязью между процессами развития и фенотипической эволюцией, а точнее — проблемой гомологии.А в последние годы, подпитываемые доступностью новых технологий и данных, вопросы, связанные с ролью процессов развития в эволюции, привлекли беспрецедентное внимание (хороший обзор см. В статьях в этом томе [Hox gene и Evo-Devo Symposium]. )
Во всех этих дискуссиях гомология была признана «центральным понятием всей биологии» (Wake, 1994), а также «нерешенной проблемой в биологии» (De Beer, 1971). Более того, не было недостатка в новых и не столь новых интерпретациях гомологии (см. e.g ., Hall, 1994; Бок и Кардью, 1999).
В этой статье я сосредоточусь на проблеме гомологии в развитии. Я разделю свое эссе на две части. Первая часть будет исторической. Здесь я продолжу проект Спемана по истории и критике концепции гомологии. Я кратко обрисую некоторые вклады в проблему гомологии за последние 75 лет и подчеркну довольно примечательную преемственность в том, как биологи думали о гомологии. Спеманн выделил три основных подхода к гомологии в XIX веке: идеалистический, исторический и причинно-аналитический.Здесь я буду утверждать, что все определения гомологии 20-го века также подпадают под одну из этих категорий (иногда включая комбинации категорий). Однако дальнейшие различия в рамках этих подходов представляют значительный интерес, если кто-то хочет понять роль концепции гомологии. в новой дисциплине эволюции развития.
Во второй части этого эссе я представлю анализ различных аспектов «гомологии в развитии». Концепция гомологии, как и все другие биологические концепции, имеет множество интерпретаций в зависимости от соответствующего теоретического контекста.В контексте эволюции развития концепция гомологии, с одной стороны, представляет собой проблему поиска ответа, а с другой стороны, концептуальный инструмент, который позволяет нам объединять различные данные (генетические, эмбриологические и сравнительные). Я буду утверждать, что именно через призму развития мы можем надеяться прийти к механистическому или биологическому пониманию гомологии. Процессы развития опосредуют разные уровни исторической гомологии, например, между гомологией генов и морфологических признаков.В каждом случае отношение гомологии устанавливается с помощью независимых критериев сравнения. И именно нетривиальное сопоставление генов и фенотипов требует механистического понимания морфологических трансформаций, эволюции и, наконец, гомологии.
ИСТОРИЯ И КРИТИЧНОСТЬ КОНЦЕПЦИИ ГОМОЛОГИИ, 1915–1999 гг.
Некоторые статьи уже касаются конкретных аспектов истории концепции гомологии (, например, ., Spemann, 1915; Panchen, 1994, 1999; Rieppel, 1994).Большинство из этих авторов, однако, сосредотачиваются на разработках XIX века (но см. Donoghue, 1992). Здесь я расскажу о нескольких вехах 20-го века, которые привели к нашей нынешней концепции гомологии. Поскольку мне придется быть избирательным, я сосредоточусь на вкладах, которые каким-то образом связаны с вопросом гомологии в развитии.
Ганс Спеманн и причинно-аналитическая концепция гомологии
По его собственному признанию, Ганс Спеманн не был сторонником теоретических рассуждений.Он предпочел детальную экспериментальную работу, чтобы установить надежные факты, которые в конечном итоге приведут к всеобъемлющему пониманию развития. Его метод был методом археолога, который «воссоздает образ бога из фрагментов, которые только он держит в руках. Он должен верить в существование целого, даже если он этого не знает; но он также не может просто воссоздать его в соответствии со своими собственными идеями. … Прежде всего он обязан уважать переломы ( Bruchflächen. ). Только тогда он может надеяться разместить новые находки в нужном месте (Spemann, 1936, стр.275) ». Спеманн использовал ту же археологическую чувствительность, когда написал свою «Историю и критику концепции гомологии» в 1915 году. В этой статье он проследил истоки современной концепции гомологии. Спеманн проводил различие между периодом идеалистической морфологии (Кампер, Гете, Жоффруа де Сен-Илер и Оуэн), историческим периодом сравнительной анатомии и филогении (Дарвин, Геккель, Гегенбаур, Мюллер) и, наконец, причинно-аналитическим периодом причинно-следственных связей. морфология и Entwicklungsmechanik.
Спеманн утверждал, что основные принципы концепции гомологии были установлены уже в идеалистический период морфологии. Гомология была основана на геометрической концепции идеального архетипа и предполагала сравнение одинаковых частей разных животных независимо от их функции. Эта версия концепции гомологии нашла свое каноническое выражение в определении гомологии Оуэном: «ГОМОЛОГ: один и тот же орган у разных животных при всех формах и функциях (Owen, 1843).Оуэн далее различал общую гомологию, сходство между морфологическим признаком и его представлением в архетипе, от особой гомологии, сходства между одним и тем же признаком у двух видов (Owen, 1848). Это различие, отождествляющее «общее» с «идеальным», было особенно характерно для идеалистического периода морфологии.
Следующий исторический период сравнительной анатомии добавляет два временных измерения к дискуссиям о гомологии: одно онтогенетическое, а другое филогенетическое.В последующие десятилетия после Дарвина специалисты по сравнительной анатомии (особенно в Германии) в основном занимались производством филогенетических деревьев. Гомология сыграла важную роль в этом стремлении. Сходство морфологических структур может быть объяснено общим происхождением, и, соответственно, идентификация гомологов может стать важным инструментом для получения филогенетических отношений. Однако это потребовало независимых критериев для идентификации гомологий. Если раньше идеальный архетип служил опорной рамкой для установления гомологии, то теперь многие исследователи обратились к эмбриологии, чтобы сделать эту работу.Эволюционные морфологи, такие как Геккель и Гегенбаур, утверждали, что истинная гомология может существовать только между двумя частями, которые развились из одного и того же зачатка (Haeckel, 1866; Gegenbaur, 1878). Интерпретация индивидуального развития (онтогенеза) как повторения филогении затем привела Геккеля к дальнейшему различению палингенеза и ценогенеза, чтобы учесть укороченные или иным образом измененные онтогенетические последовательности (Haeckel, 1866). Однако попытки найти объяснение гомологии между взрослыми признаками на ранних эмбриональных стадиях ( anlagen ) вскоре натолкнулись на трудности.
Гибкость развития, и особенно феномен возрождения, были главными эмпирическими вызовами этому преформистскому подходу к гомологии. Среди наиболее известных примеров — явления индукции и регенерации хрусталика позвоночных. При нормальном развитии хрусталик формируется из эктодермы головы в месте контакта с зрительным пузырем переднего мозга. После хирургического удаления хрусталика или даже более крупных частей глаза хрусталик регенерируется у некоторых видов земноводных.Но он больше не формируется из исходной ткани (эктодермы головы), а скорее из другого источника, дорсального края радужной оболочки. Более того, сам Спеманн обнаружил, что у некоторых видов амфибий пересаженные части эпидермиса, которые не были частью зачатка хрусталика, также могут быть индуцированы для образования хрусталика.
Чтобы противостоять этим проблемам, Спеманн повторно ввел первоначальное различие Ланкестера между гомогенией и гомоплазией (Lankester, 1870, p.39). В определении Ланкестера гомогенность относится к тем аспектам гомологии, которые можно проследить непосредственно до общего предка. Это более ограниченное определение, поскольку оно требует постоянного присутствия всех черт конкретного персонажа между видами-предками и видами-потомками. Следовательно, только более общие аспекты устройства организма будут однородными для разных видов, в то время как дальнейшие дифференциации или независимые разработки не подпадут под эту категорию. С другой стороны, в определении Ланкестера гомоплазия относится к сходству, которое возникает, «когда идентичные или почти одинаковые силы или среды действуют на две или более части организма, которые в точности или почти одинаковы.Тогда «результирующие модификации различных частей будут в точности или почти одинаковыми» (Lankester, 1870, стр. 39). Для Спемана это различие действует в том смысле, что фокусирует наше внимание на тех «силах или средах», которые являются механистической причиной органического сходства. Если смотреть таким образом, линзы, которые происходят по-разному и из разных материалов, все еще можно рассматривать как гомологичные, как и признаки двух эмбрионов / взрослых, которые развиваются из клеток, разделенных после первых делений, и в этом случае снижение гомологии до общего anlagen достигает своего предела.Концепция гомологии исторического периода (homogeny sensu Lankester) с ее упором на историческую преемственность (как онтогенетически, так и филогенетически) распадается, потому что она не может объяснить специфические особенности развития. Следовательно, утверждал Спеманн, к гомологии следует подходить из причинно-аналитического анализа развития, как это отражено в первоначальном определении гомоплазии Ланкестера.
Адольф Ремане: Систематический учет критериев гомологии
За десятилетия между двумя мировыми войнами возник дискурс «теоретической биологии», в основном в Германии, России, Нидерландах и Англии.Вопросы, связанные с концептуальными основами биологии, и здесь особенно морфологии, а также вопросы, связанные с новыми направлениями физиологии развития и генетики, были в центре этих дебатов. Концепция гомологии также обсуждалась несколько раз; однако эти трактовки гомологии в основном включали концептуальный анализ и вопросы определения. И Людвиг фон Берталанфи, и Адольф Мейер пытались различать разные формы гомологии, такие как типологическая, онтогенетико-типологическая, филогенетическая и физиологическая гомология развития (Meyer, 1926; Bertalanffy, 1934).Но в целом, хотя за эти годы был достигнут значительный прогресс в анализе концептуальных проблем, связанных с гомологией, мало что было сделано с точки зрения разработки программ оперативных исследований.
Ситуация изменилась, когда в 1952 году Адольф Ремане опубликовал свой трактат Die Grundlagen des natürlichen Systems, der vergleichenden Anatomie und der Phylogenetik (Принципы естественной системы, сравнительной анатомии и филогенетики) (Remane, 1952). Здесь Ремане обсуждал гомологию в контексте филогении и систематики.Он дает следующее обоснование своего теоретического анализа. В то время он работал над всеобъемлющим обзором всех типов животных, который позже стал основой его успешного учебника по систематической зоологии (Ремане, 1975). Поэтому важно было прояснить теоретические основы как филогении, так и систематики. Для Ремане это было возможно только благодаря глубокому пониманию принципов сравнительной анатомии, что, в свою очередь, повлекло за собой оперативное рассмотрение гомологии.
Как мы видели ранее, естественные системы классификации всегда основывались на некотором понятии сходства. В контексте филогении естественная система подразумевает, что систематические группы различаются общими признаками, которые они унаследовали от своего последнего общего предка, , то есть ., По историческим гомологам. Проблема в том, как эти общие персонажи могут быть идентифицированы в практике сравнительной анатомии. С этой целью Ремане разработал набор критериев, которые предоставили морфологу контрольный список для установления сходства, i.е. , гомология.
Критерии Ремане (три основных и три вспомогательных критерия) приводят к вероятностному аргументу в пользу гомологии. Если эти критерии выполнены, то более вероятно, что два символа гомологичны, чем что они полностью независимы. Гомология вероятна, когда есть сходство между относительными положениями персонажей в рамках общего структурного плана, сходство в структурных деталях между этими персонажами и когда существуют переходные формы. В тех случаях, когда рассматриваемые признаки слишком просты и не имеют достаточного количества структурных деталей для непосредственного сравнения по трем основным критериям, они все равно могут считаться гомологичными, если они присутствуют в большом количестве родственных видов и если есть другие подобные персонажи, имеющие аналогичное распределение.Однако вероятность гомологии снижается, если такие признаки также присутствуют у неродственных видов.
Как и следовало ожидать от того, кто пытается создать филогенетическую систему, подход Ремана к гомологии является историческим. Распределение гомологов между разными таксонами рассматривается как следствие их филогенетического родства. Но это оставляет перед лицом эпистемологической проблемы, как можно идентифицировать те гомологи, которые используются для установления филогенетических отношений между таксонами независимо от ранее существовавшей филогении.Критерии гомологии Ремане предназначены для решения этой проблемы. Но Ремане не останавливается на достигнутом. Во второй части своей книги, которая в настоящее время в основном игнорируется, он обсуждает различные «филогенетические законы», такие как биогенетический закон, принцип сохранения более ранних стадий развития, а также различные принципы дифференциации и специализации, которые могут объяснить удивительные упорядоченные преобразования, наблюдаемые в филогении. Все эти идеи помещают его прямо в историческую традицию сравнительной анатомии, описанную Спеманом.
Вилли Хенниг: Гомология в филогенетической систематике
В 1950-х годах Вилли Хенниг также начал разрабатывать свой метод филогенетической систематики (кладистики) (Hennig, 1950; Hennig, 1966). В значительной степени опираясь на богатую немецкую традицию систематики, а также на концептуальные дискуссии в теоретической биологии, Хенниг разработал рабочее определение гомологии в контексте филогенетической систематики. Хенниг исходил из предположения, что «эволюция — это трансформация формы и поведения организма» (Zimmermann, 1953).«Этот процесс органической трансформации включает анагенез, а также кладогенез. Хенниг предположил, что можно реконструировать этот процесс, , то есть ., Филогению, проследив последовательность преобразований определенных признаков. Основная идея Хеннига заключалась в том, чтобы охарактеризовать каждую новую линию преемственности одним или несколькими преобразованными персонажами, так называемыми синапоморфиями. Их следует отличать от тех состояний характера, которые являются общими для разных линий, симплезиоморфий.Таким образом, то, является ли конкретный признак или состояние признака синапоморфией или симплезиоморфией, зависит от ранга анализируемых таксонов. Но в любом случае и синапоморфии, и симплезиоморфии относятся к тождеству между персонажами. Однако они более инклюзивны, чем традиционная гомология, потому что отсутствие признака также является законным состоянием признака в филогенетическом анализе.
Методы Хеннига в конечном итоге изменили систематику, особенно после того, как в 1966 году был опубликован английский перевод его книги по филогенетической систематике (Hennig, 1966).Но хотя его методы работают в том смысле, что они позволяют логически последовательную реконструкцию филогении, они по-прежнему зависят от предварительной оценки сходства признаков, то есть . Гомологии. Хеннинг определил гомологию, чтобы включить все преобразованные состояния персонажа. Однако это определение все еще требует независимых критериев для оценки гомологии. Хенниг использовал множество методов для идентификации гомологов, таких как палеонтологические свидетельства, но он также в значительной степени полагался на Ремане, чьи критерии гомологии затем стали вспомогательными критериями в контексте определения Хеннига.
Я довольно подробно описал вклад Ремана и Хеннига в гомологию, хотя они не делают много ссылок на развитие, потому что их идеи стали основой современной филогенетической систематики. И как указали Паула Маби (2000) и другие (например, Вагнер, 1999), правильная филогения по-прежнему является основой для всей дальнейшей работы по эволюции развития.
Гэвин де Бир: возвращение к развитию
В начале 1970-х годов Гэвин де Бир вновь связал вопрос о гомологии со старыми проблемами эмбриологии (De Beer, 1971).В широко читаемом учебнике, написанном для студентов, де Бир обсудил несколько все еще нерешенных проблем гомологии. Неудивительно, что большинство этих проблем было связано с вопросами развития. Де Бир осознал, что принцип общего происхождения не решает всех трудностей, связанных с гомологией морфологических признаков. В то время как обычное происхождение может, по крайней мере, указывать на причину некоторых очевидных странностей в развитии, таких как расположение гортанного нерва у млекопитающих, который проходит назад, огибает артериальный проток, а затем бежит вперед, чтобы иннервировать гортань, указывая на гомологию артериального протока млекопитающих с 6-й артериальной дугой рыбы и гомологии гортанного нерва с 4-й жаберной ветвью блуждающего нерва, он не может дать механистического объяснения этому консерватизму эволюции.А для многих других особенностей общее происхождение даже не дает такого объяснения.
Де Бир, анализируя вклад эмбриологии и генетики в понимание проблемы гомологии, поднял несколько важных вопросов. Например, он указал, что соответствие между ранними стадиями развития не обязательно для того, чтобы взрослые признаки были гомологичными, что различные процессы индукции организатора могут приводить к гомологичным взрослым структурам, и что идентичность генов не гарантирует гомологию между признаками.Де Бир, по сути, повторил позицию Спемана от 1915 года. Поступая так, он вернул развитие дискуссии о гомологии.
Руперт Ридл: Порядок гомологии и системная теория эволюции
Через несколько лет после того, как де Бир определил гомологию как большую нерешенную проблему в биологии, Руперт Ридл представил свою «системную теорию эволюции» (Riedl, 1975; Riedl, 1978). Ридль попытался объяснить, как уже предполагает название его книги, часто удивительные проявления «порядка» в живом мире.Морфологии животных сгруппированы, и морфопространство всех возможных форм жизни в основном пусто (Gould, 1977, 1989). Эти факты нелегко согласовать с идеей постепенной эволюции, которая лежала в основе моделей популяционной и количественной генетики в то время. В начале семидесятых многие теоретики оспаривали каноническую неодарвинистскую картину эволюции, которая возникла в результате современного синтеза 1930-х и 1940-х годов (, например, ., Eldredge and Gould, 1972; Gould and Vrba, 1982; Margulis, 1982; Мейнард-Смит, и др. ., 1985). Действительно, недавно появившееся внимание к роли процессов развития в эволюции, романтическая фаза «evo-DevO» также расцвела в те годы (Wagner et al ., 2000).
Гомология занимает центральное место в теории Ридла, вероятно, в большей степени, чем в большинстве других предложений того времени. Для Ридля гомология является наиболее заметным выражением естественного порядка. В каком-то смысле гомология — это просто следствие общего происхождения. Однако филогенетические отношения объясняют только распределение гомологов, но не их механистическую причину.Но и модели количественной и популяционной генетики не могли объяснить замечательное выражение биологического порядка, о чем свидетельствует распределение гомологов. По мнению Ридля, ответ на проблему гомологии можно найти только в системных условиях развития. Их можно рассматривать как выражение четвертого порядка нормы, иерархии, взаимозависимости и традиции. В теоретической системе Ридля все четыре выражения порядка — норма, иерархия, взаимозависимость и традиция — являются частью объяснения гомологии.Гомологи рассматриваются как идентичные, то есть нормативные части, идентичность которых поддерживается системными (функциональными) взаимозависимостями внутри процессов развития, которые их порождают, и которые образуют иерархическую систему, являющуюся следствием традиции ( т. Е. ., Наследование и общее спуск). Еще одна концепция, которую Ридл ввел для описания механистических причин идентичности гомологов, — это понятие бремени. Бремя — это мера степени системной интеграции определенных персонажей в процесс развития.Чем более интегрирован персонаж в развитии, тем выше его бремя и тем стабильнее персонаж. Идея бремени тесно связана с понятием ограничений развития, которое было предложено примерно в то же время. Обе концепции признают внутренние ограничения, которые система развития накладывает на степени вариации конкретного персонажа. Они отличаются тем, что ограничения развития больше сосредоточены на ограничениях, налагаемых на вариации, тогда как бремя определяется как количественное измерение стоимости изменения персонажа, который функционально встроен в сложную развивающуюся систему взаимозависимостей.Но поскольку гомологию можно рассматривать как утверждение об ограничениях вариабельности конкретных признаков (Wagner, 1999), понятия бремени и ограничений развития стоят в начале недавнего интереса к механистическому объяснению гомологии, которая является ядром. концепции биологической гомологии (см. ниже).
Подход Ридла к проблеме гомологии содержит многие элементы более ранних концепций. Его различение между различными формами гомологии и его опора на критерии гомологии Ремане помещают его прямо в традиции сравнительной анатомии и теоретической биологии.Однако его акцент на причинном механизме объяснения гомологии делает его также наследником причинно-аналитического подхода Спемана к гомологии.
Последние десятилетия: концепция биологической гомологии, иерархическая гомология и частичная гомология
В последние годы проблеме гомологии уделяется больше внимания, чем когда-либо прежде в этом столетии. Здесь я не могу подробно обсуждать все различные предложения (но см. Donoghue, 1992; Hall, 1994; Bock and Cardew, 1999, где содержатся прекрасные обзоры текущего состояния дискуссий).Многие из новых предложений касаются разработки. Развитие играет важную роль, особенно в концепции биологической гомологии (Roth, 1984, 1988; Wagner, 1989, 1994, 1995, 1996; Donoghue, 1992). Здесь я следую изложению Вагнера (1999) основных принципов концепции биологической гомологии. Основное предположение концепции биологической гомологии состоит в том, что гомологи являются единицами фенотипической эволюции. Как таковые, они представляют собой индивидуализированные квазиавтономные части организма, которые разделяют определенные элементы и вариативные свойства.Следовательно, если два персонажа должны быть гомологичными, они могут различаться только в тех аспектах своей структуры, которые не подвержены общим ограничениям в развитии. Роль механизмов развития состоит в том, чтобы гарантировать идентичность двух структур, поскольку они ограничивают вариационные свойства квазиавтономных единиц. Ниже я подробнее исследую связи между механизмами развития и проблемой гомологии. Здесь я просто хочу упомянуть два дополнительных измерения гомологии, которые недавно привлекли некоторое внимание, — иерархическую и частичную гомологию.
Гомология может возникать между объектами на разных уровнях биологической иерархии (Riedl, 1975; Abouheif, 1997). Гомология на этих разных уровнях обычно распознается с помощью независимых критериев сравнения на каждом уровне, таких как критерии Ремана или сравнения последовательностей (см. Также Laubichler, 1999). Это открытый вопрос, в какой степени эти разные формы гомологии совпадают, , т.е. , можем ли мы вывести морфологическую гомологию на основе установленных генетических гомологов (см. Также ниже).Важное место в этом контексте занимают процессы развития. Они являются посредниками между различными уровнями гомологов (генетическими и морфологическими). Цель причинно-аналитического подхода к гомологии — найти объяснение существования морфологических гомологов в процессах развития, которые их производят. До сих пор все исследования, которые явно рассматривали этот вопрос, а не просто предполагали, что предшествующие в развитии объекты определяют статус производных персонажей, предостерегали от этой формы преформизма (см. e.г ., Wagner, 1989; Дикинсон, 1995; Abouheif, 1997).
С вопросом об иерархической гомологии связана проблема частичной гомологии (Wake, 1999). Частичная гомология предполагает, что в случае некоторых сложных символов не все элементы должны быть гомологичными, но что можно идентифицировать определенные части, которые являются гомологичными. Есть, например, случаи, такие как парные придатки гнатостомов, которые разделяют определенные механизмы развития (формирование переднезаднего паттерна), но отличаются другими (скелетогенез) (Wagner, 1999).Эти случаи обычно представляют собой иерархию общих производных символов (Hennig, 1966), которую можно интерпретировать как все более всеобъемлющий набор частичных гомологов (см. Также Lankester, 1870).
В заключение, эта краткая история концепции гомологии в 20 веке говорит нам, что гомология действительно является «центральной концепцией биологии». Мы также видели, что все вклады в гомологию попадают в три категории, обсужденные Спеманом в 1915 году. Несмотря на то, что Спеман уже сформулировал основные принципы причинно-аналитического подхода к гомологии, его программа в действительности была реализована лишь около 25 лет назад. когда новые концептуальные идеи начали бросать вызов господству современного синтеза в эволюционной биологии.Эта задача совпала с прорывами в генетике развития и привела к нынешней программе эволюции развития. В каком-то смысле круг замкнулся. К счастью, мы также можем опираться на идеи, полученные в контексте других подходов к гомологии, особенно в рамках филогенетической систематики.
ГОМОЛОГИЯ В РАЗВИТИИ
В предыдущих разделах мы видели, что различные понятия гомологии подчеркивали либо историческое, либо причинное объяснение органического сходства.Более того, стало ясно, что мы должны различать историческое или причинное (или их сочетание) объяснение органического сходства и различные критерии для установления этого сходства (см. Также Bolker and Raff, 1996). Как и в случае с большинством биологических концепций, интерпретация гомологии зависит от конкретного эталонного процесса (Laubichler, 1999; Wagner and Laubichler, 2000). В случае гомологии релевантными эталонными процессами являются эволюция и развитие. Оба являются сложными иерархическими процессами, которые связаны между собой различными способами.Поэтому неудивительно, что в зависимости от того, какой аспект эволюции или развития изучается, уместны разные интерпретации гомологии. Попытки устранить эту врожденную многомерность концепции гомологии и разработать все более «точные» определения гомологии, следовательно, довольно бесплодны. Дэвид Уэйк недавно предложил прекратить беспокоиться о том, что «такое» гомология, и начать решать интересные эмпирические вопросы, связанные с понятием гомологии, такие как застой, модульность, сохранение дизайна или скрытая гомология (Wake, 1999).Но многомерность концепции гомологии также является одной из основных причин, почему она действительно является «центральной концепцией всей биологии (Wake, 1994)». Гомология больше не привязана исключительно к морфологическим признакам; скорее, это понятие применимо ко всему, от генов до поведения (см. недавний обзор в Bock and Cardew, 1999). Повсеместное присутствие концепции гомологии в различных областях биологии делает еще более актуальным тот факт, что мы четко понимаем соответствующий референтный процесс в каждом конкретном случае.В случае гомологии последовательностей эталонный процесс молекулярной эволюции включает механизмы замены пар оснований, транслокацию хромосомных элементов, дублирование генов и т. Д. С другой стороны, эталонный процесс фенотипической эволюции, который является основой гомологии. морфологических признаков включает в себя другой набор эволюционных механизмов, таких как аллометрический рост, изменения времени событий развития, ограничения развития, канализация, модульность и т. д.
Здесь я не могу исследовать все измерения гомологии; скорее я ограничу свой анализ различными связями, существующими между гомологией морфологических признаков и процессами развития.
Существует по крайней мере четыре различных связи между морфологической гомологией и развитием. Процессы развития могут быть использованы для установления гомологии между морфологическими признаками, процессы развития могут дать объяснение гомологии между морфологическими признаками, пути развития сами могут быть гомологами, а процессы развития являются частью объяснения эволюционных инноваций, которые также можно интерпретировать как зарождающиеся гомологи.Ниже я кратко обрисую предположения и проблемы, связанные с этими четырьмя измерениями гомологии в развитии. Однако одно предостережение относится ко всем четырем случаям. Ни один из этих случаев не является «однозначным» в том смысле, что мы можем установить a priori общий набор правил, как процессы развития связаны с гомологией. Скорее, как показывают новые данные, гомология является системным свойством развивающихся систем развития. Только когда мы поймем свойства этих систем, мы сможем хоть как-то приблизиться к разгадке этой «нерешенной проблемы биологии» (De Beer, 1971).”
Процессы развития и оценка гомологии
Установление гомологии между морфологическими признаками имеет центральное значение для понимания эволюционных преобразований и инноваций. Следовательно, в течение последних 150 лет были предложены различные критерии для определения гомологии. Однако в последние годы многие исследователи склонны предполагать, что данные о последовательности и экспрессии генов преобладают над всеми другими формами доказательств при рассмотрении гомологии между морфологическими признаками ( e.g ., Quiring et al ., 1994; Halder и др. ., 1995). Это отношение напоминает более ранние попытки установить гомологию между морфологическими признаками, сравнивая их эмбриологическое происхождение ( anlagen ). Но в то время как прежний акцент на сходстве эмбриологического происхождения обычно ограничивал число возможных гомологов, нынешнее внимание к паттернам экспрессии с большей вероятностью увеличит охват гомологических отношений, тем самым сделав их неинформативными.Недавняя дискуссия о возможной гомологии между глазами позвоночных и насекомых из-за общего главного контрольного гена ( Pax-6 ) является лишь одним примером этой тенденции (Quiring et al ., 1994; Halder et al ., 1995; Даль и др. ., 1997).
Как генетический, так и эмбриологический подходы связывают морфологическую гомологию с наличием определенных элементов в онтогенетической последовательности, которая приводит к этим признакам. Это изначально преформистское представление.Проблема с таким подходом к гомологии состоит в том, что, хотя он и является привлекательным для сведения проблемы установления гомологии на морфологическом уровне к более простому вопросу о наличии или отсутствии определенных идентифицируемых элементов, в большинстве случаев он не адекватно представляет сложности процессов развития, которые создают общие сходства между гомологами. Та же роль в развитии ортологичных генов не гарантирует идентичности морфологических признаков, равно как и одинаковые пути развития не требуются для создания гомологичных признаков.Как показывают наши данные о множественной роли многих факторов транскрипции, таких как distalless , один и тот же регуляторный модуль может использоваться в разных путях развития (Panganiban et al ., 1997). Точно так же регенерированный хрусталик развивается по другому пути развития (см. Выше).
Все эти проблемы не означают, что данные последовательности и экспрессии генов бесполезны для оценки отношений гомологии между морфологическими признаками. Напротив, они предоставляют нам дополнительную информацию, которую необходимо взвешивать вместе со всеми другими формами доказательств, в отличие от систематики, где для решения вопросов филогении все чаще используются как молекулярные, так и морфологические данные.Однако мы можем заключить, что морфологическая идентичность — это системное свойство, которое необходимо понимать в контексте эталонного процесса морфологической эволюции и эволюции, а не преформистской концепции.
Процессы развития и объяснение гомологии
Целью причинно-аналитического (биологического) подхода к гомологии является механистическое объяснение феномена органического сходства. Соответствующий эталонный процесс для морфологической гомологии — эволюция развития; развития, потому что это непосредственная причина морфологических признаков и эволюции, потому что она имеет дело с органическими преобразованиями и стабильностью.Недавно Гюнтер Вагнер (1999) обрисовал в общих чертах программу исследований для эмпирической проверки концепции биологической гомологии. Его основные шаги включают выявление предполагаемых гомологов, определение надлежащей филогении, описание внутри- и межвидовых паттернов изменчивости, анализ способов развития каждого предполагаемого гомолога и проверку того, влияют ли различия в способах развития на различия в вариационных свойствах (Wagner, 1999). ).
Предположение, лежащее в основе этого подхода, состоит в том, что объяснение стабильности гомологов можно найти в свойствах процессов развития, которые их создают.Стабильность морфологических признаков подразумевает, что потенциальные вариации этих признаков ограничены или, по крайней мере, ограничены определенными способами. Это может быть выполнено с помощью ограничений, действующих на морфогенетические механизмы, а также с помощью морфостатических механизмов, которые поддерживают или стабилизируют идентичность характера (Wagner and Misof, 1993). Таким образом, вопрос гомологии связан со смежными проблемами модульности и канализации (Wagner, 1996; Wagner and Altenberg, 1996; Wake, 1999). Таким образом, мы можем заключить, что в контексте концепции биологической гомологии механистическое объяснение гомологии морфологических признаков должно включать системные свойства процессов развития как морфогенеза, так и морфостаза.
Процессы развития и эволюционные инновации
Процессы развития также были причастны к происхождению эволюционных новшеств или инноваций (Müller and Wagner, 1991, 1996; Müller and Newmann, 1999). Эволюционные инновации — зарождающиеся гомологи. Это апоморфии, составляющие основу филогенетической систематики (Hennig, 1966). В контексте эталонного процесса эволюции развития новинки представляют для нас самую большую проблему, но они также являются окном, через которое мы можем изучать роль процессов развития в формировании морфологических преобразований.
Недавно Мюллер и Ньюман (1999) предположили, что могут быть разные фазы в происхождении и становлении морфологических новшеств. Они утверждали, что эпигенетические процессы, такие как взаимодействия между клетками, тканями и окружающей средой, а также основные биомеханические свойства этих частей играют важную роль в создании и интеграции новых структур. На втором этапе эти зарождающиеся гомологи затем будут интегрированы, как генетически, так и в плане развития, чтобы функционировать как автономные организаторы конструкции организма (Müller and Newmann, 1999).Эта гипотеза предполагает важную роль процессов развития в создании органического разнообразия.
Процессы развития как гомологи
Описание процессов развития в настоящее время обычно включает характеристики регуляторных генных сетей. Недавно стало ясно, что регуляторные генные сети представляют собой модульные структуры (Wagner, 1996; Abouheif, 1999; Wake, 1999). Как модульные структуры они могут быть признаны отдельным уровнем гомологии в биологической иерархии (Riedl, 1975; Abouheif, 1997).Есть несколько важных вопросов, которые связаны с потенциальной гомологией регуляторных генных сетей. С одной стороны, нам нужно определить критерии для оценки гомологии между различными сетями. Это требует дальнейшего детального изучения взаимодействий между элементами этих сетей (структуры этих персонажей). Осложнения могут возникнуть из-за генетической избыточности. Кроме того, эти сети приобретают дополнительные регуляторные связи и новые роли в развитии в ходе эволюции (Abouheif, 1997; Gerhart and Kirschner, 1997).Хотя это не меняет основную модульную структуру сетей, затрудняет определение точных границ этих символов. Поэтому следует ожидать, что во многих случаях мы обнаружим частичную гомологию между сетями (и истинную гомологию между некоторыми элементами этих сетей). Многие компоненты очень старые, о чем свидетельствует их замечательная сохранность в разных типах (De Robertis, 1994). ) и тот факт, что они часто рекомбинируются с другими модулями для формирования новых сетей (Gerhart and Kirschner, 1997).Поэтому нам нужна хорошая филогения, прежде чем мы сможем оценить гомологию между различными сетями. Сосредоточение внимания на гомологии между регуляторными сетями также поднимает вопрос, в какой степени их гомология подразумевает гомологию морфологических признаков. Как мы видели, это старая проблема. Но из-за модульной организации биологических систем и имеющихся свидетельств множественных функций многих ключевых элементов, а также рекомбинации между элементами этих сетей, однозначные случаи будут довольно редкими.
ЗАКЛЮЧЕНИЕ
Гомология — одно из центральных понятий эволюции развития. Ожидаемый экспоненциальный рост доступных данных об экспрессии генов и последовательностях только увеличивает потребность в оперативном подходе к гомологии (Wagner, 1999). Это также подчеркивает тот факт, что не существует единой концепции гомологии, которая охватывала бы все интересные эмпирические вопросы, связанные с биологическим порядком (Riedl, 1975) или феноменом органического сходства (Wake, 1999).Причинно-аналитический подход к гомологии, , то есть , «гомология в развитии» имеет давнюю традицию, восходящую, по крайней мере, к Гансу Спеману. Механистическое или биологическое объяснение причин гомологии будет ключом к пониманию трансформации органических форм, , то есть , фенотипической эволюции (, например, ., Müller and Wagner, 1996; Raff, 1996).
Успех причинно-аналитического подхода к гомологии зависит от наличия надежных филогений (Wagner, 1999; Paula Mabee, 2000).Исторический подход к проблеме гомологии лежит в основе развития современной филогенетической систематики (кладистики). Поэтому различные подходы к проблеме гомологии сходятся в контексте эволюции развития.
Таким образом, мы могли бы заключить с помощью Спеманна (1915, стр. 63): «Существуют концепции такой центральности, что их происхождение, изменение и распад, короче говоря, их история отражает развитие науки, частью которой они являются. Гомология — такое понятие для сравнительной анатомии.(И, мы могли бы добавить, для эволюции развития.)
Я хотел бы поблагодарить организаторов за приглашение принять участие в этом симпозиуме, а также Гюнтера Вагнера и Ханса-Йорга Райнбергера за проницательные обсуждения по теме этой статьи и двух анонимных рецензентам за полезные комментарии.
Список литературы
Abouheif
,
E.
1997
. Генетика развития и гомология: иерархический подход.
Тенденции в экологии и эволюции
,
12
405
-408.
Abouheif
,
E.
1999
. Установление критериев гомологии для регуляторных генных сетей: перспективы и проблемы. In Г. Р. Бок и Г. Кардью (ред.), Стр. 207–222 Гомология. Симпозиум Фонда Новартис 222, Вили, Чичестер.
Bertalanffy
,
L. v.
1934
. Wesen und Geschichte des Homologiebegriffes.
Unsere Welt
,
28
.
Бок
,
г.R.
и G. Cardew (ред.).
1999
. Гомология. Симпозиум Фонда Новартис 222. John Wiley & Sons, Чичестер.
Bolker
,
J. A.
и R. A. Raff.
1966
. Генетика развития и традиционная гомология.
BioEssays
,
16
489
-494.
Даля
,
E.
и Х. Косеки. и др.
1997
. Гены Pax и органогенез.
BioEssays
,
19
755
-765.
Де Бир
,
г.
1971
. Гомология, нерешенная проблема . Издательство Оксфордского университета, Оксфорд.
Де Робертис
,
Э. М.
1994
. Гомеобокс в дифференцировке и эволюции клеток. В D. Duboule (ed.), Guidebok к генам гомеобокса. Издательство Оксфордского университета, Оксфорд.
Дикинсон
,
W. J.
1995
. Молекулы и морфология: где гомология?
Тенденции в генетике
,
11
119
-121.
Донохью
,
М. Дж.
1992
. Гомология. В Э. Ф. Келлер и Э. А. Ллойд (ред.), Ключевые слова эволюционной биологии . Издательство Гарвардского университета, Кембридж, Массачусетс.
Eldredge
,
N.
и S. J. Gould.
1972
. Прерывистые равновесия: альтернатива филетическому градуализму. В Т. Дж. М. Шопф (ред.), Модели в палеобиологии . Фриман, Сан-Франциско.
Gegenbaur,
1878
. Grundriß der vergleichenden Anatomie. . Энгельманн, Лейпциг.
Gerhart
,
J.
и M. Kirschner.
1997
. Клетки, эмбрионы и эволюция. . Блэквелл, Молден, Массачусетс.
Gilbert
,
S. F.
и J. M. Opitz. и др.
1996
. Ресинтезирование биологии развития.
Разработка. Биол
,
173
357
-372.
Гулд
,
С.J.
1977
. Онтогенез и филогения. . Издательство Гарвардского университета, Кембридж, Массачусетс.
Гулд
,
С. Дж.
1989
. Прекрасная жизнь. . W. W. Norton. Нью-Йорк.
Gould
,
S. J.
и E. Vrba.
1982
. Exaptation: отсутствующий термин в науке о форме.
Палеобиология
,
8
4
-15.
Haeckel
,
E.
1866
. Generelle Morphologie der Organismen. . Реймер, Берлин.
Гальдер
,
Г.
и П. Каллаертс. и др.
1995
. Индукция эктопических глаз путем направленной экспрессии гена eyeless у дрозофилы . .
Science
,
267
1788
-1792.
Холл
,
Б. К.
1992
. Эволюционная биология развития.. Чепмен и Холл, Лондон.
Холл
,
Б. К.
(ред.)
1994
. Гомология. Иерархическая основа сравнительной биологии. . Academic Press, Сан-Диего.
Хенниг
,
W.
1950
. Grundzüge einer Theorie der Phylogenetischen Systematik. . Берлин, Deutscher Zentralverlag.
Хенниг
,
W.
1966
. Филогенетическая систематика. .Издательство Иллинойсского университета, Урбана.
Ланкестер
,
р.
1870
. Об использовании термина гомология.
Летопись и журнал естествознания, зоологии, ботаники и геологии
,
6
34
-43.
Лаубихлер
,
М. Д.
1999
. Семиотический взгляд на биологические объекты и биологические функции.
Семиотика
,
127
415
-431.
Маби
,
П. М.
2000
. Данные о развитии и филогенетическая систематическая эволюция конечностей позвоночных.
амер. Зоол
,
40
789
-800.
Маргулис
,
Л.
1982
. Симбиоз в клеточной эволюции. . Фриман, Сан-Франциско.
Maynard-Smith
,
J.
и R. Burian. и др.
1985
. Ограничения развития и эволюция.
кварт. Ред. Биол
,
60
265
-287.
Мейер
,
А.
1926
. Logik der Morphologie im Rahmen einer Logik der gesamten Biologie. . Юлиус Спрингер, Берлин.
Müller
,
G. B.
и S. A. Newmann.
1999
. Генерация, интеграция и автономия: три шага в эволюции гомологии. В Г. Р. Бок и Г. Кардью (ред.), Гомология. Симпозиум Фонда Новартис 222, стр.65–72. Уайли, Чичестер.
Müller
,
G. B.
и G. P. Wagner.
1991
. Новинка в эволюции: реструктуризация концепции.
Annu. Rev. Ecol. Syst
,
22
229
-256.
Müller
,
G. B.
и G. P. Wagner.
1996
. Гомология, Hox генов и интеграция в процессе развития.
амер. Зоол
,
36
4
-13.
Оуэн
,
Р.
1843
. Лекции по сравнительной анатомии и физиологии беспозвоночных животных, прочитанные в Королевской коллегии хирургов в 1843 году. . Лонгман, Браун, Грин и Лонгманс, Лондон.
Оуэн
,
Р.
1848
. Об архетипе и гомологиях скелета позвоночных. . Джон ван Ворст, Лондон.
Panchen
,
A. L.
1994
. Ричард Оуэн и концепция гомологии. В Б. К. Холл. (ред.), Гомологии. Иерархическая основа сравнительной биологии, с. 22–62. Academic Press, Сан-Диего.
Panchen
,
A. L.
1999
. Гомология — история понятия. В Г. Р. Бок и Г. Кардью (ред.), Гомология. Симпозиум Фонда Новартис 222, стр. 5–17. Джон Вили и сыновья, Чичестер.
Panganiban
,
G.
и S. M. Irvine. и др.
1997
.Происхождение и эволюция придатков животных.
Proc. Natl. Акад. Sci. США
,
94
5162
-5166.
Quiring
,
R.
et. al
1994
. Гомология гена безглазых дрозофилы с геном малых глаз у мышей и аниридии у людей.
Science
,
265
785
-789.
Рафф
,
Р.А.
1996
. Форма жизни. Гены, развитие и эволюция формы животных.Издательство Чикагского университета, Чикаго.
Ремане
,
А.
1952
. Die Grundlagen des natürlichen Systems, der vergleichenden Anatomie und der Phylogenetik. . Akademische Verlagsgesellschaft, Лейпциг.
Remane
,
A.
и V. Storch. и др.
1975
. Systematische Zoologie. . Густав Фишер, Штутгарт.
Ридл
,
р.
1975
. Die Ordnung des Lebendigen. . Парей, Берлин и Гамбург.
Ридл
,
р.
1978
. Порядок в живых организмах. . John Wiley & Sons, Нью-Йорк.
Риппель
,
О.
1994
. Гомология, топология и типология: история современных дебатов. В Б. К. Холл (ред.), Гомология. Иерархическая основа сравнительной биологии, с. 63-100. Academic Press, Сан-Диего.
Roth
,
V. L.
1984
. О гомологии.
Biol. J. Linn. Soc
,
22
13
-29.
Roth
,
V. L.
1988
. Биологические основы гомологии. В К. Дж. Хамфрис (ред.), Онтогенез и систематика, стр. 1-26. Издательство Колумбийского университета, Нью-Йорк.
Spemann
,
H.
1915
. Zur Geschichte und Kritik des Begriffs der Homologie. In C. Chun и W. Johannsen (ред.), Allgemeine Biologie. B. G. Teubner, Leipzig und Berlin
3
.4.1
63
-85.
Spemann
,
H.
1936
. Experimentelle Beiträge zu einer Theorie der Entwicklung. . Спрингер, Берлин.
Вагнер
,
Г. П.
1989
. Концепция биологической гомологии.
Ann. Rev. Ecol. Syst
,
20
51
-69.
Вагнер
,
Г. П.
1994
. Гомология и механизмы развития. В Б. К. Холл (ред.), Гомология: иерархическая основа сравнительной биологии, стр. 273–299 Academic Press, Сан-Диего.
Вагнер
,
Г. П.
1995
. Биологическая роль гомологов: гипотеза строительных блоков.
N. Jb. Геол. Paläont. Abh
,
195
279
-288.
Вагнер
,
г.П.
1996
. Гомологи, естественные виды и эволюция модульности.
амер. Зоол
,
36
36
-43.
Вагнер
,
Г. П.
1999
. Программа исследований для проверки концепции биологической гомологии. In G. Bock and G. Cardew (ред.), Homology (Novartis Foundations Symposium 222) , стр. 125–134. Уайли, Чичестер.
Вагнер
,
Г. П.
и Л. Альтенберг.
1996
. Сложные адаптации и эволюция эволюционируемости.
Evolution
,
50
967
-976.
Wagner
,
G. P.
и M. D. Laubichler.
2000
. Идентификация характера в эволюционной биологии: роль организма.
Теория в биологических науках
,
119
20
-40.
Wagner
,
G. P.
и B. Y. Misof.
1993
. Как можно ограничить развитие персонажа, несмотря на вариации в путях развития?
Дж.Эволюционная биология
,
6
449
-455.
Wagner
,
G. P.
, C-h. Чиу и М. Лаубихлер.
2000
. Эволюция развития как механистическая наука :. Вывод от механизмов развития к эволюционным процессам
амер. Зоол
,
40
819
-831.
Пробуждение
,
D.
1999
. Гомоплазия, гомология и проблема «тождества» в биологии. In G. Bock and G. Cardew (ред.), Homology (Novartis Foundations Symposium 222), стр. 24–33. Уайли, Чичестер.
Пробуждение
,
Д. Б.
1994
. Сравнительная терминология.
Science
,
265
268
-269.
Zimmermann
,
W.
1953
. Evolution. Geschichte ihrer Probleme und Erkenntnisse. . Альбер, Фрайбург и Мюнхен.
Общество интегративной и сравнительной биологии
Icon 3 — Гомология | Национальный центр естественнонаучного образования
Гомология
Гомология — это конкретное объяснение сходства форм, наблюдаемых в биологическом мире.Сходства часто можно объяснить общим происхождением; признаки считаются гомологичными, если показано, что они унаследованы от общего предка. Например, хотя руки четырехконечных позвоночных выглядят внешне совершенно по-разному, все они имеют одинаковый базовый скелетный и мышечный паттерн. Такие общие шаблоны лучше всего объясняются выводом о том, что они унаследованы от общего предка, у которого также был этот шаблон. Предлагаемые гомологии оцениваются с использованием сравнительной анатомии, генетики, развития и поведения.
Уэллс едет на карусели гомологии …
Уэллс утверждает, что гомология используется биологами циклически, потому что учебники определяют гомологию как сходство, унаследованное от общего предка, а затем заявляют, что гомология является доказательством общего предка . Уэллс прав: это упрощенное прочтение гомологии действительно круглое. Но Уэллс излишне упрощает сложную систему до абсурда, вместо того, чтобы пытаться объяснить ее должным образом. Уэллс, как некоторые биологи и многие учебники, допускает классическую ошибку, смешивая определение гомологии с диагнозом гомологичной структуры, биологическую основу гомологии с процедурой открытия гомологии.В своем обсуждении он путает не только природу концепции, но и ее историю; в результате возникает дискуссия, которая сбивает с толку. Здесь действительно важно не то, описывают ли учебники гомологию по кругу, а то, используется ли гомология в биологии по кругу. Когда гомология правильно понимается и применяется, она совсем не круговая.
… назад в будущее
До 1859 года, когда Дарвин опубликовал Происхождение видов , гомология определялась как сходство структуры и положения и отличалась (хотя и непоследовательно) от «аналогии», которая определялась как сходство функций, но не обязательно структуры и положения.Примером гомологии и аналогии являются крылья птиц и летучих мышей. Руки птиц и летучих мышей будут считаться гомологами, потому что они имеют одинаковую структуру и положение у обоих животных. Их крылья, однако, являются аналогами. Оба крыла выполняют одну и ту же функцию (полет), но крыло птицы сделано из перьев, а крыло летучей мыши — из кожи. Это разные конструкции.
Додарвиновской основой сходства была идея «архетипа». Однако архетип так и не получил четкого определения.Идея принадлежит морфологической теории, которая пришла от немецких философов-трансценденталистов конца 1700-х — начала 1800-х годов. Это было в значительной степени вышло из моды к 1840-м годам, но Ричард Оуэн, который систематизировал это различие, был привержен философии трансцендентных причин, как отмечали многие историки науки (например, Russell 1916 ; Desmond 1982 ; Rupke 1993 ; Padian 1995a , 1997 ).
Тем не менее, структура биологического мира больше похожа на генеалогию, чем на галерею архетипов «печенья».«Дарвин объяснил сходство в строении и положении очень разных животных результатом естественного отбора, работающего над общими предковыми моделями. Концепция гомологии сместилась от отражения нечеткого« архетипа »к отражению происхождения с модификацией. по-прежнему диагностируют гомологичные структуры, сначала ища структуры сходной формы и положения, как это делали додарвиновские биологи (они также ищут генетические, гистологические, эволюционные и поведенческие сходства.) Однако в наш постдарвиновский период биологи определяют гомологичную структуру как анатомическую, онтогенетическую, поведенческую или генетическую особенность, разделяемую между двумя разными организмами, потому что они унаследовали ее от общего предка. Поскольку не все признаки, сходные у двух организмов, обязательно унаследованы от общего предка, и не все признаки, унаследованные от общего предка, похожи, необходимо проверить структуры , прежде чем их можно будет объявить гомологичными.Чтобы ответить на вопрос, «может ли эта особенность в этих группах быть унаследована от общего предка?» ученые сравнивают этот признак во многих группах, ища закономерности формы, функции, развития, биохимии, а также наличия и отсутствия. Многие признаки одновременно проверяются на соответствие генеалогии посредством процесса, который Клюге ( 1997 ; см. Также Kluge, 1998 , 1999 для обсуждения независимых тестов на гомологию) назвал проверкой «множественных специальных гипотез гомологии».»
Если с учетом всех имеющихся свидетельств распределение характеристик по множеству различных групп напоминает генеалогический образец, весьма вероятно, что этот признак отражает общее происхождение. Однако будущие тесты, основанные на большем количестве характеристик и большем количестве групп, могут изменить эти оценки, однако — что является нормальным для построения научного понимания. Тем не менее, когда очень большой объем информации из нескольких различных областей (анатомия, биохимия, генетика и т. Д.) Указывает на то, что набор организмов имеет генеалогическое родство, тогда ученые чувствуют себя уверенно, заявляя общие черты гомологичны.Наконец, хотя суждения о гомологии в принципе можно пересмотреть, во многих случаях нет реалистичных ожиданий, что они будут отменены.
Итак, Уэллс ошибается, когда говорит, что гомология предполагает общее происхождение. Отражает ли признак общее происхождение двух или более групп животных — протестировано против образца , который он создает с этими, а также с другими группами. Иногда, хотя и не всегда, образец отражает генеалогические отношения между организмами, и в этот момент делается вывод об общем происхождении.Сегодня процесс тестирования осуществляется методом, называемым «филогенетическая систематика» или «кладистика», который может выполняться без предположения об эволюционной взаимосвязи между группами, но происхождение с модификацией является лучшим объяснением закономерностей, которые выявляет сравнение признаков. .
Эволюция и гомология — тесно связанные концепции, но они не являются круговыми: гомология структуры диагностирована и проверена внешними элементами: структурой, положением и т. Д., и является ли образец распределения признака генеалогическим. Если модель взаимоотношений выглядит как генеалогия, было бы неправильно отрицать, что эта черта отражает общее происхождение или что между группами существуют эволюционные отношения. Точно так же близость отношений между двумя группами организмов определяется степенью гомологичных признаков; чем более гомологичны два организма, тем более недавний их общий предок. Вопреки утверждению Уэллса, ни определение, ни применение гомологии к биологии не являются круговыми.
Как уже упоминалось, новые данные из различных областей биологии расширили наше понимание гомологии за пределы анатомических структур. Анатомические гомологии, поведенческие гомологии, гомологии развития и генетические гомологии могут быть независимо диагностированы и протестированы.
Поведенческая гомология распознает особенности поведения животных, которые могут быть прослежены до общего предка. Например, рассмотрим способы гнездования птиц и крокодилов. Обе эти группы разделяют строение гнезд, родительскую заботу о детенышах и «пение» для защиты территории и привлечения партнеров.Большинство людей знают, что птицы делают эти вещи, но меньше людей знают, что их кузены, аллигаторы и крокодилы, тоже делают то же самое. Они унаследовали это поведение от общего предка. Из-за гомологии мы предполагаем такое поведение и для их вымерших предков; поэтому неудивительно, что окаменелости многих нептичьих динозавров были найдены гнездящимися со своими детенышами ( Хорнер и Макела, 1979, ; , Хорнер, 1982, ; , Кларк и др., 1999, ).
Гомологии развития — это особенности программ развития организмов.Примером этого являются «глоточные мешочки», которые почти все позвоночные в той или иной степени приобретают в процессе своего развития, но которые становятся совершенно разными структурами у взрослых особей. Например, эмбриологические глоточные карманы бесчелюстных хордовых (например, Amphioxus , микс и миноги) развиваются в глоточные дуги и щели, которые поддерживают структуру жабр и позволяют воде выходить из глотки после прохождения через жабры. У челюстных позвоночных, таких как акулы и рыбы, глоточные мешки развиваются в жаберные опоры и части скелета челюсти.У наземных позвоночных (четвероногих) эти дуги и карманы развиваются в скелет челюсти и мускулатуру, но другие карманы / дуги, которые у жаберных позвоночных развились в структуру жабр, теперь развиваются в кости и полости уха, а также в хрящи щитовидной железы и трахеи ( Gilbert 2000). ). Эволюция различных морфологий глотки взрослых позвоночных является результатом изменений этих эмбриональных структур и их компонентов посредством программы развития ( Graham 2001 ).
Сегодня мы также распознаем генетические гомологии. Есть похожие гены, которые контролируют развитие негомологичных признаков. Например, есть ген под названием « Pax6 », которым обладают плодовые мушки, мыши и многие другие организмы, который влияет на развитие глаза. Биологи предполагают, что ген унаследован от общего предка не только из-за его биохимического сходства, но и из-за его распространения во многих таксонах. Однако настоящие глаза, которые образуют эти гены, не являются результатом общего происхождения — их общий предок, скорее всего, не обладал глазами, хотя, возможно, он обладал светочувствительной способностью.Глаза мух, мышей и многих других существ имеют различное строение и положение и исторически не непрерывны, однако ген Pax6 исторически непрерывен и отвечает за них всех. Этот гомологичный ген функционирует как «переключатель», который запускает развитие светочувствительных органов Gilbert 2000 , но запускаемые ими гены «ниже по течению» уже не те же самые: они управляют разными программами развития и, таким образом, строят структурно разные глаза в мухи, мыши и другие организмы.Относительно новая область эволюционной биологии развития (сокращенно evo-DevO) занимается этими процессами. Открытия, сделанные всего за последние 10 лет в этой области, значительно расширили наше понимание гомологии и сделали картину более сложной. Уэллс почти игнорирует эту важную новую область в своем обсуждении, что является удивительным упущением для человека, чей опыт включает в себя степень в области биологии.
Гомология, эволюция и природа науки
Некоторые формулировки концепции гомологии кажутся круговыми, но, как обсуждалось выше, существует внешний референт (образец , который характеристики принимают в разных группах), который служит Независимый тест, концепция, правильно определенная и понятая, не является.Утверждение Уэллса о круговороте гомологии раскрывает ошибочное представление о том, как работает наука. В науке идеи часто формулируются путем перехода между данными и теорией, и ученые регулярно проводят различие между определением концепции и доказательствами, используемыми для ее диагностики и проверки.
Гомология на самом деле не более круговая, чем методы, используемые в геологии для определения палеогеографии и тектоники плит. Например, в 1920-х годах Альфред Вегенер использовал форму континентов, корреляцию слоев горных пород, корреляцию ископаемых организмов и положение ледниковых полос в качестве доказательства своего предположения о том, что континенты когда-то были объединены в один суперконтинент и имели впоследствии «дрейфовали» к своим текущим местоположениям.Сегодня геологи могут оценить, где раньше находилась определенная часть континента, глядя на полярные блуждания, палеомагнетизм, ледниковые полосы, корреляцию слоев и окаменелостей и форму. Является ли это более замкнутым, чем рассуждение о гомологии? Нет. Доказательства были использованы для вывода о том, что континенты перемещались, а затем концепция тектоники плит была применена к различным данным для определения положения континентов в разное время. Аналогия с тектоникой плит также имеет отношение к выводу Уэллса ( Wells 2000 : 77) о том, что мы не полностью понимаем механизмы гомологии: механизм распространения морского дна, возможно, еще не полностью понят, но континенты все еще движутся.
Что говорят учебники
Представления о гомологии в учебниках, рассмотренных Уэллсом, различаются только продолжительностью их обсуждений. В целом учебники содержат 2–10 абзацев (обычно включая обсуждение аналогий и рудиментарных признаков). Поскольку в более коротких вводных учебниках мало места для изучения сложностей определения, диагностики и применения гомологии, их объяснения граничат с циркуляром. В более длинных учебниках высшего уровня проводится более четкое различие между объяснением гомологии (общее происхождение) и использованием наборов гомологий для восстановления отношений (рис. 7).Все учебники включают схемы передних конечностей различных позвоночных и все гомологические элементы, кроме одного, для облегчения идентификации. Гутман включает в себя второй рисунок, показывающий гомологичные кости у нескольких четвероногих и один череп рыбы, наглядно демонстрирующий, как были изменены черепа. Футуйма, Гутман, Кэмпбелл и др. включать лучшие обсуждения и иллюстрации гомологии, но, тем не менее, получить D от Уэллса.
Большинство учебников включают обсуждения аналогий и рудиментарных структур наряду с обсуждениями гомологии.Аналогичные признаки — это особенности со схожими функциями (но не обязательно схожими структурами), которые не унаследованы от общего предка, но эволюционировали конвергентно, тогда как рудиментарные признаки — это остаточные структуры, которые были сохранены от предыдущих форм. Примечательно, что Уэллс упускает из своей оценки любое упоминание об аналогичных особенностях или рудиментарных структурах (например, поясах конечностей у змей или поясах конечностей китов, цитируемых в большинстве учебников).
Оценка по учебнику Уэллса
Согласно Уэллсу, учебники должны объяснять, что гомологии — это сходства структуры и функций не из-за общего происхождения, а из-за общего «архетипа» или базового плана, на котором основывались все формы (Уэллс удивительно уклончив в том, что касается что он имеет в виду под «архетипом»).Когда Уэллс предлагает вернуться к додарвиновскому взгляду на гомологию в учебниках, он не объясняет, что это будет значить для преподавания биологии или биологии. Он не объясняет, что это заменит проверяемую модель (происхождение) несуществующей, непроверяемой трансценденталистской конструкцией. Уэллс расплывчат, потому что он просто хочет продвинуть свою позицию, а архетип согласуется с некоторым понятием особого творения, которое поддерживают сторонники «разумного замысла» креационизма и их союзники.
Оценки Уэллса (он ставит только D и F), по-видимому, коррелируют с продолжительностью охвата учебником (рис. 7).Например, все книги с оценкой D посвящают обсуждению гомологии более 200 слов, тогда как три книги с оценкой F посвящают менее 200 слов. Это потому, что разница между D и F для Уэллса заключается в том, определяет ли книга гомологию «циклически». Следовательно, способность «хорошо» относиться к гомологии (что означает D) во многом зависит от того, сколько места отведено для ее обсуждения. Однако Уэллс позволяет в книге иметь изображение. Чтобы получить оценку B или выше, учебники должны определять гомологию как сходство структуры и положения и утверждать, что гомология основана на концепции «архетипа».Кроме того, они должны заявить, что «архетип» может означать многое, а не только общее происхождение. Он также хочет, чтобы в учебниках было указано — неточно — что такие механизмы, как генетика и программы развития, не учитывают гомологию. Наконец, он хочет, чтобы учебники учитывали гомологию. заявляют, что концепция гомологии «противоречива». Эта схема сфальсифицирована на провал, потому что современная биология не считает гомологию противоречивой или основанной на архетипах. Конечно, нет никаких оснований принимать эти критерии классификации.
Почему мы все еще должны учить, что гомология является результатом общего происхождения
Как показывают наши нынешние знания биологии, нет причин сомневаться в том, что паттерны структур, поведения, генов и программ развития лучше всего соответствуют гипотеза о том, что все организмы имеют общих предков. Многие сходства между этими сильно расходящимися группами являются результатом их происхождения. В биологии в настоящее время обсуждают не вопрос о том, реальна ли гомология, а скорее, какие структуры гомологичны и как мы можем наилучшим образом определить гомологию (поскольку наши диагностические подходы ошибочны).Такое обсуждение надежности методологии характерно для всех областей науки, а не только для биологии. Происхождение — основа гомологии; похожие гены, действуя через развитие, передают гомологию между поколениями. Гены строят структуры посредством своего взаимодействия в программе развития. Следовательно, гены, развитие и сходство структуры и положения — это процедуры обнаружения гомологии; они помогают биологам определять эволюционные отношения. Это соответствует образцам и процессам, которые мы наблюдаем в мире природы; вот чему мы должны учить.
Как в учебниках можно улучшить обсуждение гомологии
Самый большой недостаток в описании гомологии в учебниках состоит в том, что они, как и Уэллс, имеют тенденцию путать определение гомологии с диагностикой гомологичных признаков. В учебниках необходимо прямо указать, что гомологии — это сходства, наблюдаемые в биологическом мире, которые лучше всего объясняются общим происхождением. Затем они должны объяснить, как диагностируются гомологичные структуры по их сходной структуре и положению, биохимической основе, пути развития и так далее.Более подробное и расширенное обсуждение поможет устранить видимость округлости, вызванную чрезмерно упрощенными описаниями. Описание того, как гомология используется в качестве инструмента для открытия эволюционных взаимосвязей, сделало бы ее гораздо более педагогически полезной концепцией для студентов, поскольку она показала бы им, как эволюционные биологи используют анатомические наблюдения для обнаружения эволюционных взаимосвязей. Наконец, добавление понятий нескольких уровней гомологии из генетики и биологии развития лучше покажет студентам, как сходятся разные линии доказательств, чтобы поддержать гомологии и филогении.Учебники не должны следовать предложению Уэллса о том, что гомология — это просто сходство по структуре и положению, а также не должны указывать, что существуют «другие» причины гомологий, помимо наследования от общего предка. Если вернуться к понятию гомологии Уэллса 19-го века, студенты окажутся неподготовленными к участию в науке 21-го века.
Эволюционная генетика
3. Эволюция понимает гомологии
Ричард Оуэн (1848) ввел термин гомология для обозначения структурного сходства между организмами.
Для Оуэна это сходство указывало на то, что организмы были созданы по общему плану или архетипу.
То есть, хотя каждый вид уникален, планы для каждого могут иметь много общих черт, так же как планы дизайна для Honda Civic и Honda Prelude могут быть похожими.
Тем не менее, если бы каждый организм был создан независимо, непонятно, почему среди одних организмов было бы так много гомологий, а среди других — так мало.
Также трудно понять тот факт, что гомологичные структуры могут быть неэффективными или даже бесполезными.
Почему у некоторых пещерных рыб были выродившиеся глаза, которые не могли видеть?
Дарвин придал смысл гомологичным структурам, предоставив им эволюционное объяснение:
- У родственных организмов сходная структура, потому что все эти организмы произошли от общего предка, у которого был эквивалентный признак.
Ридли использует особое определение гомологии: «Сходство между видами, которое не является функционально необходимым».
Я интерпретирую это так: «Сходство между видами существует, несмотря на несколько вероятных альтернативных черт, которые могли бы функционировать одинаково хорошо.»
3. Эволюция понимает гомологии
«Универсальный» генетический код
| 5 мин. | т | С | A | G | 3 ‘ |
|---|---|---|---|---|---|
| т | TTT Phe (F) TTC Phe (F) TTA лей (L) TTG лей (л) | TCT Ser (S) TCC Ser (S) TCA Ser (S) TCG Ser (S) | ТАТ Тюр (г) TAC Tyr (Y) TAA Стоп TAG Stop | TGT Cys (C) ТГК Цис (К) TGA Stop [ Trp (W) ] TGG Trp (Вт) | т С А г |
| С | CTT Leu (L) CTC Leu (L) CTA лей (L) CTG Leu (L) | CCT Pro (P) CCC Pro (P) CCA Pro (P) CCG Pro (П) | Кошка His (H) CAC His (H) CAA Gln (Q) CAG Gln (Q) | CGT Arg (R) CGC Arg (R) CGA Arg (R) CGG Arg (R) | т С А г |
| А | ATT Ile (I) УВД Иле (И) ATA Ile (I) [ Met (M) ] ATG Met (M) | ACT Thr (T) ACC Thr (T) ACA Thr (T) АЧГ Thr (Т) | AAT Asn (N) AAC Asn (N) AAA Lys (К) AAG Lys (K) | АГТ Сер (S) AGC Ser (S) AGA Arg (R) [ Stop ] AGG Arg (R) [ Stop ] | т С А г |
| G | GTT Val (В) GTC Val (В) GTA Val (В) ГТГ Вал (В) | ГКТ Ала (А) GCC Ala (A) GCA Ala (A) GCG Ala (А) | GAT Asp (D) GAC Asp (D) GAA Glu (E) GAG Glu (E) | GGT Gly (G) GGC Gly (G) GGA Gly (G) GGG Gly (G) | т С А г |
[ Курсивом обозначен митохондриальный код млекопитающих.]
Генетический код генов, кодирующих белок, почти универсален у эукариот и прокариот.
Исключения составляют большинство митохондриальных геномов и некоторые ядерные (например, Mycoplasma и Tetrahymena).
Даже в этих случаях генетический код очень похож.
Существуют миллионы альтернативных генетических кодов, так почему же у всех организмов почти один и тот же?
Поскольку антикодон находится на противоположном конце от сайта связывания аминокислоты тРНК и не взаимодействует с сайтом связывания, нет химической необходимости в назначении кодона конкретной аминокислоте.
Генетический код у живых организмов гомологичен: он похож, несмотря на то, что существует много одинаково хороших генетических кодов.
Однако при гипотезе о произошедшей эволюции сходство всех генетических кодов имеет смысл:
- Общий предок всех известных организмов имел генетический код, аналогичный тому, что мы видим сегодня.
На протяжении веков генетический код передавался без изменений (или почти так) от родителей к потомству, потому что мутации генетического кода были бы катастрофическими (изменение аминокислотной последовательности всех продуцируемых белков).
(Что бы подумал эволюционист, если бы сегодня был обнаружен организм с совершенно другим генетическим кодом?)
3. Эволюция понимает гомологии
Плазменная мембрана
Точно так же плазматические мембраны всех организмов, эукариотических и прокариотических, структурно похожи и состоят из двухслойного фосфолипида.
Существует много других возможных мембранных структур.
Хвосты гидрофобных жирных кислот могут быть соединены. Может быть три цепи гидрофобных жирных кислот.Помимо глицеринфосфорной кислоты, могут быть задействованы и другие гидрофильные группы.
Сходство плазматической мембраны (а также других клеточных структур) предполагает, что все живые клетки произошли от предка со сходной структурой мембраны.
3. Эволюция понимает гомологии
Пентадактильная конечность
Одним из классических примеров гомологичной структуры является пентадактильная (= пятизначная) конечность.
Все четвероногие (= четвероногие) имеют конечности с пятью пальцами, по крайней мере, на некоторой стадии развития.
Некоторые четвероногие теряют часть этих пальцев в процессе развития, как показано на изображении крыла птицы.
Но если крылу птицы не нужны пять пальцев, почему у растущего зародыша изначально пять пальцев развиваются?
Наиболее правдоподобное объяснение состоит в том, что, хотя пять цифр не являются функционально необходимыми, они представляют собой генетический артефакт, унаследованный от предков птиц.
3. Эволюция понимает гомологии
Новые функции развиваются из уже существующих структур
Гомологические структуры преподают нам важный урок эволюции:
- Evolution работает в основном за счет изменения уже существующих структур.
То есть, даже когда два вида функционируют совершенно по-разному, они часто используют гомологичные структуры для выполнения этих функций.
Например, птицы и летучие мыши скорее летают, чем бегают на четвереньках, но их крылья представляют собой видоизмененные передние конечности, а не совершенно новую структуру.
Точно так же жало ос и пчел — это модифицированный яйцеклад, а не совершенно новая структура. (Объясняя, почему жалит только самки пчел !!)
3. Эволюция понимает гомологии
Остатки
Структуры, которые не имеют функции у одного вида, но гомологичны функционирующей структуре у других видов, особенно трудно объяснить, кроме как в рамках теории общего происхождения.
Такие структуры известны как рудиментарные структуры.
Например:
Тазовый пояс кита
Глазные луковицы слепых пещерных существ, таких как грот-саламандра (Typhlotriton spelaeus):
Пыльники и пыльца бесполых одуванчиков:
Рудиментарные структуры крайне нелогичны, если каждое существо было создано независимо, но имеют смысл, если организмы наследуют черты своих предков с постепенными изменениями с течением времени.
4. Гомологии вложены
Одной из поразительных особенностей подобных структур является то, что они группируются.
Определенные виды имеют много общего на всех уровнях организации (например, люди и шимпанзе), тогда как другие виды имеют только определенные почти универсальные гомологии (например, люди и Escherichia coli).
Это наблюдение не имеет особого смысла для созданных объектов, поскольку создатель может смешивать и сочетать особенности, наблюдаемые в любом организме.
Напротив, согласно теории эволюции, мы ожидаем, что количество общих гомологий будет высоким для близкородственных видов и уменьшаться со временем по мере того, как виды расходятся друг от друга.
Рассмотрим последовательность ДНК четырех видов:
Здесь виды C и D будут иметь идентичные последовательности, виды A и B будут отличаться только одной мутацией, тогда как A-C и A-D будут отличаться четырьмя мутациями, а B-C и B-D — тремя мутациями.
Фактически, «родство» между организмами определяется тем, сколько (и какие) общие черты они имеют.
4. Гомологии вложены
Примат Филогения
Хотя разные черты могут немного по-разному отражать эволюционную историю группы видов, должно быть приблизительное соответствие.
Например, морфологически люди и шимпанзе гораздо больше похожи, чем люди и гиббоны.
Если это связано с тем, что люди и шимпанзе более тесно связаны (= недавно у них был общий предок), то можно ожидать, что последовательности ДНК человека и шимпанзе также будут более похожими.
Три филогенетических дерева были реконструированы на основе последовательностей ДНК:
- (а) 4700 п.н. митохондриальной ДНК,
(б) семенниковоспецифический белок на Y-хромосоме,
(c) некодирующие области гена -глобина.
Все три дерева показывают, что последовательности ДНК человека и шимпанзе в среднем более похожи, чем последовательности человека и гиббона или человека и орангутанга.
4. Гомологии вложены
Филогения птиц
Гомологические признаки также могут быть идентифицированы в ископаемых организмах, где они также имеют тенденцию проявлять вложенные серии, а не неперекрывающиеся категории.
Среди окаменелостей динозавров некоторые таксоны имеют несколько общих черт с птицами, в то время как другие имеют несколько общих черт.
В частности, манирапторы имеют больше общих черт с современными птицами, чем любой другой тип динозавров.
Точно так же целурозавры, помимо манирапторов, имеют некоторые общие черты, но не такие, как у птиц.
Считается, что эта вложенная серия представляет собой эволюционную траекторию динозавров по линии происхождения, которая привела к птицам.
Вот некоторые черты, которые классифицируют различные группы архозавров (включая крокодилов, динозавров и птиц):
Архозаврия
- Яйца с твердой скорлупой
- одиночные отверстия на каждой стороне черепа перед глазами (анторбитальные отверстия)
- отверстия в нижней челюсти (нижнечелюстное отверстие)
- высокий узкий череп с заостренной мордой
- набор зубьев в гнездах
Динозаврия
- уменьшенные четвертые и пятые цифры на руке
- стопа уменьшена до трех основных пальцев
- Три или более позвонка, составляющих крестец
- открытая тазобедренная розетка
Saurischia
- хватательная рука
- асимметричные пальцы
- длинная подвижная шея
- Таз с лобковой костью, направленной вниз и вперед
Therapoda
- кости полые тонкостенные
Целурозаврия
- удлиненные руки
- щиколотки с развитой петлей
- (у некоторых целурозавров есть открытые, как у птиц, гнезда, в которых вынашивались яйца)
Maniraptora
- полулунный запястный сустав на запястье
- модифицированная передняя конечность (приподнятая передняя конечность с возможностью складывания)
- Сращение ключицы и грудины
- Таз с направленным вниз лобком
Авес
- перья
- крылья
- поперечный рычаг
- большой палец ноги противопоставления
Самая ранняя из известных ископаемых птиц (археоптерикс) разделяет эти черты, но также имеет многие черты терапевтов, в том числе:
- зубьев
- обширные хвостовые позвонки
- когти
- Отсутствие килевидной грудки
Вероятно, археоптерикс не летал.Ему не хватало выдающегося киля современных птиц, на котором крепятся мускулы полета. (Возможно, это был планер.)
5. Свидетельства эволюции из летописи окаменелостей
Трилобит Эволюшн
Хотя летопись окаменелостей часто бедна и неполна, есть определенные отложения, где осадочные слои остаются почти непрерывным рядом.
Окаменелости из этих серий являются прямым свидетельством эволюционных изменений.
Шелдон (1987) исследовал серию осадочных слоев ордовикского периода (500 млн лет назад), содержащих окаменелости трилобитов (вымершие морские членистоногие).
Образцы были получены каждые три миллиона лет. Количество ребер у каждого вида трилобитов со временем менялось (= эволюция).
Некоторые из этих изменений со временем были настолько значительными, что животные в конце серии были отнесены к новому роду!
5. Свидетельства эволюции из летописи окаменелостей
Фораминифер эволюция
Еще более точный масштабный анализ был проведен Malmgren et al. (1983) по видам фораминифер (раковинных простейших) с 10МЯ до недавнего времени.
[Представлены три эпохи: миоцен (M; 23,8-5,2 млн лет назад), плиоцен (P; 5,2-1,8 млн лет назад) и плейстоцен (Q; 1,8 млн лет назад — 10 000 лет назад)].
За этот период окаменелые раковины превратились в более крупную и толстую раковину с более выраженным гребнем.
Хотя летопись окаменелостей демонстрирует, что изменение происходило непрерывно (= без разрывов и скачков), скорость изменения не всегда была одинаковой: форма изменилась больше всего на границе миоцена / плиоцена.
Эти изменения были достаточно значительными, чтобы линия была отнесена к виду Globorotalia plesiotumida в миоцене, а впоследствии к виду Globorotalia tumida .
Летопись окаменелостей показывает, что эволюционные изменения действительно происходят.
Недостаток летописи окаменелостей состоит в том, что обычно трудно определить силы отбора, которые могли способствовать этим изменениям.
Преимущество летописи окаменелостей перед современными наблюдениями за эволюцией состоит в том, что можно отслеживать эволюционные изменения более высокого порядка (например, происхождение новых видов, новых родов и т. Д.).
ИСТОЧНИКИ:
- Пентадактильные конечности: Ridley (1997) Evolution.
- Кит, саламандра, деревья приматов: Фриман и Херрон (1998) эволюционный анализ.
- Фотография мембраны: Wessells and Hopson (1988) Biology.
- Информация о динозаврах: Палеонтологический музей Калифорнийского университета.
- Летопись окаменелостей трилобитов и фораминифер: Футуйма (1998) Эволюционная биология.
Вернуться на домашнюю страницу Biology 336.
Функциональная группа и гомологическая серия | Примечания, видео, контроль качества и тесты | 11 класс> Химия> Основы органической химии
Функциональная группа и гомологичная серия
Функциональная группа
Органическое соединение состоит из двух частей: реактивной (активной) части, известной как функциональная группа, и скелетной части, состоящей из атомов углерода и водорода, которая известна как алкильная группа (ее общая формула — C n H 2n + 1 ) и обозначается -R).
Таким образом, функциональная группа определяется как атом или группа атомов, связанных с углеродом, которые определяют характерные свойства органических соединений. Химические свойства органического соединения зависят исключительно от природы функциональной группы, присутствующей в молекуле. Например —
Функциональная группа спирта -ОН.
Функциональная группа альдегида -CHO.
Функциональная группа карбоновой кислоты -COOH.
Некоторые важные функциональные группы и их классы показаны в таблице ниже —
| S.N | Функциональная группа | Название | Класс соединений | ||||||
| 1. | -O- | Эфир (алкокси) | Простые эфиры 12 | 2, | 2,912 | Спирт (гидрокси) | Спирты | ||
| 3. | -NH 2 | Амин (амино) | Амины | ||||||
| 4. | , Br, I = X (X) | Галогенид (галоген) | Алкилгалогениды | ||||||
| 5. | -CN или -C≡N | Цианид (нитрил) | Цианиды или нитрил | ||||||
| 6. | -NC или | Изоцианид | Изоцианид или карбиламины | ||||||
| 7. | -CNO или -C≡N → O | Цианат | Цианаты | ||||||
| 8. — | Изоцианат | Изоцианаты | |||||||
| 9. | -NO 2 или | Nitro | Nitro | ||||||
| 10. | -NO или -N = O | -Nitroso | 11. 90 90 | Азо | Азосоединения | ||||
| 12. | -CHO или | Альдегид (формил) | Альдегиды | ||||||
| 13. | Кето | Кетоны | |||||||
| 14. | -COOH или | Карбоксил | Карбоновая кислота | ||||||
| 15. | -COOR или | или 9122CO | 0 0 0| 0 Сложный эфир | Ацилгалогенид | Ацилгалогениды | |
Гомологический ряд
Когда органические соединения, содержащие одну и ту же функциональную группу, расположены последовательно на основе их возрастающей молекулярной массы, имеющей разность -CH 2 (метилен группа) между двумя последовательными соединениями, такая серия называется гомологической серией.Отдельные члены гомологичного ряда известны как гомологичные, а процесс известен как гомология.
Различные члены гомологического ряда могут быть обозначены общей формулой, например —
Алкан (C n H 2n + 2 ), аминогруппа (C n H 2n + 1 .NH . 2 ), спирт (C n H 2n + 1 .OH) и т.д. +2 )
| Формула | Название ИЮПАК | |
| CH 4 | Метан | |
| CH 3 -CH 9 3 91812229 -CH 2 -CH 3 | Пропан | |
| CH 3 -CH 2 -CH 2 -CH 3 | Бутан | |
| CH18 2 -CH 2 -CH 2 -CH 3 | Пентан | |
| CH 3 -CH 2 -CH 2 -CH 2 -CH 4 -CH 3 |
6 Hexane
- Гомологический ряд спиртов (C n H 2n + 1 .OH)
| Формула | Название IPUAC | |||
| CH 3 -OH | Метанол | |||
| CH 3 -CH 2 -OH690 9129 3 -CH 2 -CH 2 -OH | Пропанол | |||
| CH 3 -CH 2 -CH 2 -CH 2 -OH | CH 3 -CH 2 -CH 2 -CH 2 -CH 2 -OH | Пентанол | ||
| CH 3 -CH 2 -CH 2 -CH 2 -CH 2 -CH 2 -CH 2 -CH 2 -OH | Гексанол | |||
| CH 3 -CH 2 -CH 2 -CH 2 -CH 2 -CH 2 -CH 2 -CH 2 -OH | Гептанол | |||
| CH 9180 9 3 -CH 2 -CH 2 -CH 2 -CH 2 -CH 2 -CH 2 -CH 2 -OH | Октанол |
Характеристики гомологическая серия
1) Все члены гомологической серии имеют одинаковую общую формулу и сходные структуры,
2) Два последовательных члена серии отличаются по своей молекулярной формуле на -CH 2 единиц или 14 атомных атомов масса (a.м.у.).
3) Из-за наличия одной и той же функциональной группы все члены гомологического ряда почти одинаковы по химическим свойствам.
4) Все члены одной гомологической серии могут быть получены аналогичными общими методами приготовления.
5) Существует регулярная градация физических свойств, таких как температура плавления, плотность и т. Д. Среди членов одного и того же гомологического ряда.
6) Первый член гомологического ряда несколько отличается по химическим свойствам от последующих членов.
[ИЮПАК — Международный союз чистой и прикладной химии]
Ссылка
Адхикари, Рамешвар; Ханал, Сантош; Субба, Бимала; Адхикари, Сантош; Khatiwada, Shankar Pd. Универсальная химия XI . Первый. Vol. 1-й. Катманду: Публикация Оазиса, 2069.
Чаудхари, Ганга Рам; Карна, Шила Кант Лал; Шарма, Канчан; Сингх, Санджай; Гупта, Дипак Кумар. Учебник высшей средней химии XI .Эд. 2-й. Катманду: Видьярти Пустак Бхандар, 2069 (2012).
гомологических и аналогичных структур: в чем разница?
Вы когда-нибудь задумывались о том, что у разных видов животных части тела работают одинаково? Как летучие мыши и пчелы? У обоих существ есть крылья, хотя их тела устроены по-разному.
Ключ к пониманию этих различий заключается в знании того, что такое гомологичные и аналогичные структуры. В этой статье мы поговорим о каждом типе структуры и обсудим сходства и различия между ними.
Что такое гомологичные структуры?
Гомологические структуры аналогичны структурам у родственных организмов. Самое важное, что нужно помнить о гомологичных структурах, — это то, что они имеют общее происхождение. Другими словами, только организмы, которые так или иначе связаны друг с другом, могут иметь гомологичные структуры.
Например, рука шимпанзе и рука человека являются гомологичными структурами. Оба набора рук имеют схожую структуру и использование, а шимпанзе и люди имеют общего предка.
Что такое аналогичные конструкции?
Аналогичные структуры — это аналогичные структуры у неродственных организмов. Эти структуры похожи, потому что они выполняют одну и ту же работу, а не потому, что имеют общее происхождение.
Например, у дельфинов и акул есть плавники, хотя они не связаны между собой. У обоих видов плавники развились в зависимости от того, как (и где) они живут.
В чем разница между гомологичными и аналогичными структурами?
Гомологические и аналогичные структуры имеют несколько ключевых отличий.Посмотрим на них:
| Гомологические структуры | Аналогичные структуры |
Пример гомологичных структур
Прекрасным примером гомологичных структур являются крылья летучей мыши и руки человека. Летучие мыши и люди — млекопитающие, поэтому у них общее происхождение.
Крыло летучей мыши и человеческая рука имеют схожую внутреннюю структуру кости , хотя внешне они выглядят по-разному. Крыло и рука также выполняют разные функции. — крылья помогают летучим мышам летать, а руки помогают людям совершенно по-другому взаимодействовать со своим миром.
Пример аналогичных структур
Отличным примером аналогичных структур являются крыло летучей мыши и крыло пчелы. Летучие мыши и пчелы не имеют общего предка, поэтому структуры не могут быть гомологичными.
Крылья летучей мыши и пчелы служат одной цели — помогают летучим мышам и пчелам летать! Внешне конструкции тоже похожи.Однако их внутреннее строение сильно различается: крылья летучей мыши имеют костную структуру с мускулами, а крылья пчелы представляют собой перепончатые отростки.
Гомологические и аналогичные структуры — основные выводы
У многих животных части тела выглядят одинаково, хотя у них нет общих функций. У других животных части тела выглядят совершенно иначе, но имеют общий фон. Вот разница между гомологичными и аналогичными структурами:
- Гомологичные структуры — это структуры, которые могут выглядеть или функционировать иначе, чем у родственных организмов.
- Аналогичные структуры — это структуры, которые выглядят и функционируют одинаково у неродственных организмов.
Что дальше?
В настоящее время изучаете биологию и вам нужна помощь по другим темам, связанным с биологией? Узнайте о фотосинтезе, ферментах, клеточной теории, а также о том, что и как работают клеточная мембрана и эндоплазматический ретикулум.
Думаете ли вы о сдаче предметных тестов SAT по такому предмету, как биология? Убедитесь, что вы получили инсайдерскую информацию о тесте, прежде чем принимать решение.
Если вы первокурсник, второкурсник или ученик , вы можете подумать о добавлении классов AP в свое расписание. Вот полный список курсов и тестов AP, которые помогут вам выбрать подходящие для вас занятия.
Хотите узнать о самых быстрых и простых способах преобразования между градусами Фаренгейта и Цельсия? Мы вас прикрыли! Ознакомьтесь с нашим руководством по лучшим способам преобразования Цельсия в градусы Фаренгейта (или наоборот).
Извлечение нецелевых серий гомологов из данных масс-спектрометрии высокого разрешения с дефисом | Journal of Cheminformatics
Основываясь на определении серии сигналов LC-HRMS, которые могут быть вызваны гомологичными соединениями, последовательное обнаружение проходит в два этапа.Первый этап извлекает набор S
3 возможных трех кортежей (триплетов) пиков, в то время как второй этап поэтапно рекомбинирует их в более крупные кортежи из n > 3.
Определение серии
Серия A k длины n ≥ n
мин.
определяется как кортеж S
н, к
= ( п.
1 , к,
…, с
н, к
) выбранных пиков сигнала LC-HRMS p = { m / z, RT, интенсивность }, упорядоченных путем увеличения m / z пиков серии. S
n
обозначает набор всех таких последовательных кортежей, имеющих длину n . Предполагается, что соседние пики в кортеже отличаются только повторяющейся и, возможно, неизвестной химической единицей или функциональной группой, например, CH 2 или OH. В результате изменения разницы масс Δ m / z между любыми двумя соседними пиками серии p
к, к
и p
к +1 , к
должен оставаться в пределах погрешности [-4 ε; 4 ε ]. ε здесь означает максимальную ошибку измерения ± m / z и может зависеть от m / z или пиковой интенсивности [36]. Δ m / z для всех серий в наборе данных LC-HRMS находится в пределах нижней и верхней границ Δ m / z
мин.
и Δ m / z
макс.
, априори установленный как рассматриваемый диапазон масс химических единиц при заданных зарядах z .
Кроме того, Δ m / z ограничивает возможные различия в дефекте массы соседних пиков серии, обозначенные как Δ m . Дефект массы здесь относится к отклонению между точным значением m / z иона и его ближайшим целым числом [37]. Для любой моноизотопной химической единицы, которая может составлять разность масс Δ m / z , границы γ
мин.
и γ
макс.
для минимальной и максимальной разницы Δ м между сериями пик p
к, к
и еще один пик р
к +1 , к
можно определить по дефектам массы изотопов с наименьшей массой для каждого из элементов, содержащихся в единице.Например, хотя и не зная точного состава химического элемента, но предполагая, что присутствуют только C, H, N, O, S, Cl и Br, мы можем ожидать значение Δ м между любыми двумя пиками серии. отличается на Δ m / z , чтобы лежать в пределах [-0,0010 Δ m / z ; 0,0078 Δ m / z ]. Первый фактор γ
мин.
определяется отношением дефекта массы к атомной массе 79 Br, второй фактор γ
макс.
по отношению к 1 H.Коэффициенты для всех других элементов находятся между этими границами. γ необходимо рассчитать по всем химическим элементам, если нельзя сделать никаких предположений об участвующих элементах. Математическое определение γ
мин.
и γ
макс.
приведен в Дополнительном файле 1. Кроме того, необходимо учитывать округление, используемое при расчете массовых дефектов: любое Δ м вдоль ряда, приводящее к значениям массового дефекта выше 0.5 соответственно оборачивают их до Δm — 1, тогда как значения ниже -0,5 преобразуются в Δ м + 1. Таким образом, разницы на Δ м должны быть соответственно адаптированы.
Подобно границам для Δ m / z и Δ m , отклонения во времени удерживания RT между соседними пиками серии также должны быть ограничены, чтобы отразить разумные хроматографические характеристики, вызванные повторным введением химических единиц [29 , 30]. С одной стороны, Δ RT
мин.
и Δ RT
макс.
, следовательно, определяют минимальные и максимальные границы для различий в RT от одного пика p
к, к
до его следующего пика кортежа p
к +1 , к
соответственно.С другой стороны, можно ожидать, что изменения Δ RT в серии будут систематическими [29, 30]. Во-первых, такие изменения Δ RT от одной пары соседних пиков в кортеже ( p
к, к
, п.
к +1 , к
) к следующему ( p
к +1 , к
, п.
к +2 , к
) должно быть меньше заданного значения, обозначенного как ΔΔ RT .Во-вторых, кубические сглаживающие сплайны приспособлены к модели RT как функция m / z в каждом последовательном наборе [38]. Вкратце, соответствие модели каждой серии определяется коэффициентом детерминации ( R
2 ) должен быть выше определенного порога с использованием предварительно заданного параметра сглаживания λ ≥ 0.
Обнаружение триплетов
Ограничено указанными выше границами для Δ m / z , Δ m и ΔRT , первый этап обнаружения серии использует k -мерных ( k -d) деревьев [39] в качестве метрической структуры данных, чтобы обеспечить вычислительно быстрое извлечение пиковых трех кортежей, которые могут быть возможными подкортежами более крупных серий. кортежи.{4} \)
$$ a_ {x} = \ left ({m / z_ {x}, \, \ Delta m_ {x} — \, \ gamma_ {min} m / z_ {x}, \, \ Delta m_ {x}) } — \, \ gamma_ {max} m / z_ {x}, \, RT_ {x}} \ right) $$
(1)
Геометрическое изображение элементов в и
x
обозначен синими линиями на рис. {3} \)).Начиная с первой записи a
x
для корневого узла и рекурсивно циклически перебирая записи a
x
до тех пор, пока не будут достигнуты разделы с одним пиком (т. Е. Конечные узлы), дерево k -d поддерживает запросы быстрого диапазона. Эти запросы выполняются для извлечения всех пиков из двух подпространств типов L и H с более низким и высоким m / z , многократно центрированными на каждом пике LC-HRMS.Извлеченные пики затем последовательно рекомбинируют для отбора всех уникальных комбинаций пиков, которые могут образовывать 3 кортежа в соответствии с приведенным выше определением ряда и которые включают текущий центральный пик в качестве второго элемента триплета (ср. Зеленые и синие точки на рис. . 1). Пики, запрашиваемые с помощью этих подпространств, обычно представляют собой незначительную часть всех измеренных пиков LC-HRMS, и рекомбинация до возможных триплетов, таким образом, значительно улучшается по сравнению с проверкой с использованием всех пиков. Упомянутые типы подпространств L и H являются результатом объединения интервалов
$$ I_ {1} = [a _ {{x_ {1}}} + \ Delta m / z_ {min}; a _ {{x_ {1}}} + \ Delta m / z_ {max}] $$
(2)
$$ I_ {2} = [a _ {{x_ {2}}} — 2 \ varepsilon; \ infty] $$
(3)
$$ I_ {3} = [- \ infty; a _ {{x_ {3}}} + 2 \ varepsilon] $$
(4)
$$ I_ {4} = \ left [{a _ {{x_ {4}}} + RT_ {min}; a _ {{x_ {4}}} + RT_ {max}} \ right] $$
(5)
$$ I_ {5} = [a _ {{x_ {1}}} — \ Delta m / z_ {max}; a _ {{x_ {1}}} — \ Delta m / z_ {min}] $$
(6)
$$ I_ {6} = [- \ infty; a _ {{x_ {2}}} + 2 \ varepsilon] $$
(7)
$$ I_ {7} = \ left [{a _ {{x_ {3}}} — 2 \ varepsilon; \ infty} \ right] $$
(8)
$$ I_ {8} = [a _ {{x_ {4}}} — RT_ {max}; a _ {{x_ {4}}} — RT_ {min}] $$
(9)
через их декартовы произведения на запрошенные подпространства
$$ H = I_ {1} \ times I_ {2} \ times I_ {3} \ times I_ {4} $$
(10)
$$ L = I_ {5} \ times I_ {6} \ times I_ {7} \ times I_ {8} $$
(11)
Фиг.1
Примерный запрос подпространства ( черных многоугольников ) для обнаружения кортежей из 3-х элементов с центром на одном пике ( синяя точка ) с одним обнаруженным 3-кортежем, представленным в зеленом . Пересечения синих линий с осями обозначают значения четырех элементов и .
x
для этого пика. Кроме того, пунктирных линий обозначают интервал m / z , определенный в уравнении.(6). В свою очередь, пунктирных линий на верхней панели обозначают интервал RT уравнения. (9), тогда как пунктирных линий на нижней панели указывают верхнюю и нижнюю границу интервалов, связанных с различиями дефектов массы из уравнений. (7) и (8) соответственно
Интервалы I
1 – I
4 определяют границы в каждом из четырех измерений a
x
для подпространства H , следующего за запрошенным центральным пиком.Напротив, интервалы I
5 – I
8 определяют подпространство L , предшествующее запрошенному пику, как показано черными линиями и многоугольниками на рис. 1. Дополнительные сведения о том, как учесть упомянутую проблему округления Δ м и ускорить вычисление подпространств представлены в дополнительном файле 2.
Рекомбинация кортежей
На втором этапе последовательно извлеченные 3-кортежи объединяются в более крупные кортежи.Для этого все попарные комбинации кортежей x и y из набора S
n
, которые различаются только их первым и последним пиковыми членами, то есть
$$ \ left ({p_ {1, x}, \ ldots, \, p_ {n — 1, x}} \ right) = \ left ( {p_ {2, y}, \ ldots, \, p_ {n, y}} \ right) $$
(12)
или
$$ \ left ({p_ {2, x}, \ ldots, \, p_ {n, x}} \ right) = \ left ({p_ {1, y}, \ ldots, \, p_ {n — 1, y}} \ right) $$
(13)
и соответствуют приведенным выше определениям серии, касающимся ε , изменения в ΔRT и λ объединяются в новый (n + 1 ) — набор в S
n +1 .После формирования всех комбинаций из S
n
, результирующие кортежи в S
n +1 , в свою очередь, рекомбинируются в более крупные кортежи в следующем наборе S
n +2 . Это повторяется до тех пор, пока не будет достигнут пустой набор, при этом каждый набор n может свободно объединяться с несколькими различными наборами ( n + 1).Следовательно, пик не может быть включен более одного раза в один кортеж, но несколько раз в несколько разных кортежей. В каждой рекурсии n -элементов, которые могут быть объединены по крайней мере в один новый ( n + 1) -кортеж в S
n +1 удалены из S
n
; в противном случае они остаются в S
n
или отбрасываются, если длина меньше минимальной, определяемой пользователем n
мин.
.Более того, избыточные подкортежи, которые образуются путем регулярного пропуска пиков в больших n -кортежах наборов S
n ≥5 необходимо фильтровать при каждой рекурсии.
Спаривание серий
Как уже упоминалось, пик может быть членом более чем одной серии, поскольку содержащиеся в нем 3 кортежа могли быть включены в несколько различных более крупных кортежей вместо одного. Чтобы выяснить основные причины такой неоднозначности, все пары уникальных серий, которые пересекаются по крайней мере в одном пике образца LC-HRMS, были извлечены, и их свойства были охарактеризованы двояко.
С одной стороны, угол пересечения θ использовался для аппроксимации того, насколько две серии x и y такой пары пересекались в плоскости RT и m / z . θ определяется как
$$ \ cos \ theta = \ frac {{u_ {x} \ cdot u_ {y}}} {{\ parallel u_ {x} \ parallel \ parallel u_ {y} \ parallel} } $$
(14)
В этом уравнении числитель и знаменатель определяют скалярное произведение и произведение евклидовой нормы векторов с масштабированными средними значениями
$$ u_ {x} = \ left ({\ frac {{\ overline {{\ Delta RT_ { x}}}}} {{c _ {\ Delta RT}}}, \ frac {{\ overline {{\ Delta m / z_ {x}}}}}} {{c _ {\ Delta m / z}}}} \ справа) $$
(15)
каждой серии соответственно.Здесь \ (c _ {\ Delta RT} \) и \ (c _ {\ Delta m / z} \) — это диапазон значений \ (\ overline {\ Delta RT} \) и \ (\ overline {\ Delta m / z} \) по всем сериям. Чем меньше значение θ , тем больше две парные серии накладываются друг на друга в плоскости RT по сравнению с плоскостью m / z . При θ = 0 π они полностью перекрывают друг друга.
С другой стороны, самоорганизующаяся карта (SOM) использовалась для визуализации и кластеризации общих свойств среди парных рядов для объяснения различий в θ [40, 41].Будучи стратегией обучения без учителя, SOM позволяют отображать большой набор из м многомерных входных векторов v = ( v
1 ,…, v
j
,…, v
кв.м
) свойств последовательной пары на меньшую двумерную сетку узлов SOM. Затем SOM можно выборочно отображать для сопоставленных свойств; аналогичные свойства здесь сопоставлены с близкими областями в SOM, в то время как различные довольно разделены.В данном случае каждый входной вектор свойств пары рядов
$$ v_ {j} = \ left ({\ frac {{\ overline {{\ Delta RT_ {x}}}}}} {{\ hat {c} _ {\ Delta RT}}}, \ frac {{\ overline {{\ Delta m / z_ {x}}}}} {{\ hat {c} _ {\ Delta m / z}}}, \ frac { {\ overline {{\ Delta RT_ {y}}}}} {{\ hat {c} _ {\ Delta RT}}}, \ frac {{\ overline {{\ Delta m / z_ {y}}}} } {{\ hat {c} _ {\ Delta m / z}}}} \ right) $$
(16)
содержит средние значения разностей m / z и RT , присутствующих в парных сериях x и y , расположенных в виде \ (\ overline {{\ Delta m / z_ {x}}} \ ge \ над чертой {{\ Delta m / z_ {y}}} \) и \ (\ hat {c} _ {\ Delta RT} \) и \ (\ hat {c} _ {\ Delta m / z} \), представляющие средние ожидаемые погрешности измерения Δ RT и Δ m / z , соответственно.На основе этих свойств угол пересечения θ может быть вычислен для каждого узла SOM с помощью уравнения. (14) для оценки суперяркости отображаемых на него рядов. Дополнительная информация об обучении и качестве SOM представлена в Дополнительном файле 3. Расчеты SOM проводились с пакетом R kohonen , параметризованным, как указано в Дополнительном файле 4: Таблица S1 [42].
Отбор проб и анализ
Оценка проводилась на 24-часовых пробах, пропорциональных потоку, отобранных из сточных вод десяти швейцарских очистных сооружений в феврале 2010 года, как использовалось и подробно описано в Schymanski et al.[16]. Короче говоря, каждый объем образца 0,25 л регулировали pH, фильтровали, добавляли 103 меченых изотопами стандартов и обогащали посредством твердофазной экстракции в смешанном слое. После щелочной / кислотной экстракции, дальнейшего обогащения в потоке газообразного азота, восстановления водой для ВЭЖХ до 1 мл и второй стадии фильтрации конечную аликвоту 20 мкл л анализировали с помощью HPLC-ESI-HRMS. Хроматографическая стадия включала колонки Waters XBridge C18 (Милфорд, США) и градиент вода / метанол при скорости потока 200 мкм л / мин, создаваемый смесительным насосом низкого давления Rheos 2200 (Flux instruments, Базель, Швейцария).Q-Exactive (Thermo Fisher Scientific, Сан-Хосе, США) использовали для масс-спектрометрического анализа полного сканирования с разрешением 140000 при m / z = 200, после ионизации электрораспылением в каждом положительном и отрицательном режимах (напряжение распыления + 4 и −3 кВ соответственно; температура капилляра 350 ° C). Холостое измерение проводилось перед каждым блоком аликвот положительных и отрицательных образцов соответственно. Файлы данных находятся в открытом доступе через репозиторий MassIVE [43].
Обработка данных
Данные полного сканирования LC-HRMS были центроидированы и преобразованы в открытые файлы формата mzXML с помощью ProteoWizard (версия 3.0,7162) [44, 45]. Затем весь последующий анализ проводился в статистической среде R [46]. Используя пакет R enviPick (версия 1.2) [47], ионные хроматограммы были извлечены в каждый файл, и каждая извлеченная хроматограмма была проверена на наличие пиков сигнала с параметрами, перечисленными в дополнительном файле 5: Таблица S2. После выбора пика серии были обнаружены с помощью описанного выше алгоритма, как параметризовано в Дополнительном файле 6: Таблица S3. Для каждого пика, являющегося частью серии, вычитание пустого и деизотопирование выполнялось с помощью enviMass v3.1 [48] и nontarget v1.9 [49] пакетов, соответственно (параметры в дополнительных файлах 7 и 8 см. В таблицах S4 и S5). В первом случае для каждого пика образца проверяли центрированное по пикам окно RT и m / z , чтобы оно не содержало необработанных контрольных точек данных, превышающих интенсивность пика максимальной пробы более чем в 0,1 раза, чтобы подтвердить его присутствие в эффлюенте. Правило большинства, то есть доля ≥0,5 пиков на серию, использовалось для окончательного определения серии, которая должна иметь пустое происхождение.Для деизотопинга сравнение с квантованными данными моделирования позволило сгруппировать пики изотопологов неизвестного соединения в пределах заданной погрешности измерения.