
Содержание
Метод наименьших квадратов
Начнем статью сразу с примера. У нас есть некие экспериментальные данные о значениях двух переменных – x и y. Занесем их в таблицу.
| i=1 | i=2 | i=3 | i=4 | i=5 | |
| xi | 0 | 1 | 2 | 4 | 5 |
| yi | 2,1 | 2,4 | 2,6 | 2,8 | 3,0 |
После выравнивания получим функцию следующего вида: g(x)=x+13+1.
Мы можем аппроксимировать эти данные с помощью линейной зависимости y=ax+b, вычислив соответствующие параметры. Для этого нам нужно будет применить так называемый метод наименьших квадратов. Также потребуется сделать чертеж, чтобы проверить, какая линия будет лучше выравнивать экспериментальные данные.
В чем именно заключается МНК (метод наименьших квадратов)
Главное, что нам нужно сделать, – это найти такие коэффициенты линейной зависимости, при которых значение функции двух переменных F(a, b)=∑i=1n(yi-(axi+b))2 будет наименьшим. Иначе говоря, при определенных значениях a и b сумма квадратов отклонений представленных данных от получившейся прямой будет иметь минимальное значение. В этом и состоит смысл метода наименьших квадратов. Все, что нам надо сделать для решения примера – это найти экстремум функции двух переменных.
Иначе говоря, при определенных значениях a и b сумма квадратов отклонений представленных данных от получившейся прямой будет иметь минимальное значение. В этом и состоит смысл метода наименьших квадратов. Все, что нам надо сделать для решения примера – это найти экстремум функции двух переменных.
Как вывести формулы для вычисления коэффициентов
Для того чтобы вывести формулы для вычисления коэффициентов, нужно составить и решить систему уравнений с двумя переменными. Для этого мы вычисляем частные производные выражения F(a, b)=∑i=1n(yi-(axi+b))2 по a и b и приравниваем их к 0.
δF(a, b)δa=0δF(a, b)δb=0⇔-2∑i=1n(yi-(axi+b))xi=0-2∑i=1n(yi-(axi+b))=0⇔a∑i=1nxi2+b∑i=1nxi=∑i=1nxiyia∑i=1nxi+∑i=1nb=∑i=1nyi⇔a∑i=1nxi2+b∑i=1nxi=∑i=1nxiyia∑i=1nxi+nb=∑i=1nyi
Для решения системы уравнений можно использовать любые методы, например, подстановку или метод Крамера. В результате у нас должны получиться формулы, с помощью которых вычисляются коэффициенты по методу наименьших квадратов.
n∑i=1nxiyi-∑i=1nxi∑i=1nyin∑i=1n-∑i=1nxi2b=∑i=1nyi-a∑i=1nxin
Мы вычислили значения переменных, при который функция
F(a, b)=∑i=1n(yi-(axi+b))2 примет минимальное значение. В третьем пункте мы докажем, почему оно является именно таким.
Это и есть применение метода наименьших квадратов на практике. Его формула, которая применяется для поиска параметра a, включает в себя ∑i=1nxi, ∑i=1nyi, ∑i=1nxiyi, ∑i=1nxi2, а также параметр
n – им обозначено количество экспериментальных данных. Советуем вам вычислять каждую сумму отдельно. Значение коэффициента b вычисляется сразу после a.
Обратимся вновь к исходному примеру.
Пример 1
Здесь у нас n равен пяти. Чтобы было удобнее вычислять нужные суммы, входящие в формулы коэффициентов, заполним таблицу.
| i=1 | i=2 | i=3 | i=4 | i=5 | ∑i=15 | |
| xi | 0 | 1 | 2 | 4 | 5 | 12 |
| yi | 2,1 | 2,4 | 2,6 | 2,8 | 3 | 12,9 |
| xiyi | 0 | 2,4 | 5,2 | 11,2 | 15 | 33,8 |
| xi2 | 0 | 1 | 4 | 16 | 25 | 46 |
Решение
Четвертая строка включает в себя данные, полученные при умножении значений из второй строки на значения третьей для каждого отдельного i. Пятая строка содержит данные из второй, возведенные в квадрат. В последнем столбце приводятся суммы значений отдельных строчек.
Пятая строка содержит данные из второй, возведенные в квадрат. В последнем столбце приводятся суммы значений отдельных строчек.
Воспользуемся методом наименьших квадратов, чтобы вычислить нужные нам коэффициенты a и b. Для этого подставим нужные значения из последнего столбца и подсчитаем суммы:
n∑i=1nxiyi-∑i=1nxi∑i=1nyin∑i=1n-∑i=1nxi2b=∑i=1nyi-a∑i=1nxin⇒a=5·33,8-12·12,95·46-122b=12,9-a·125⇒a≈0,165b≈2,184
У нас получилось, что нужная аппроксимирующая прямая будет выглядеть как y=0,165x+2,184. Теперь нам надо определить, какая линия будет лучше аппроксимировать данные – g(x)=x+13+1 или 0,165x+2,184. Произведем оценку с помощью метода наименьших квадратов.
Чтобы вычислить погрешность, нам надо найти суммы квадратов отклонений данных от прямых σ1=∑i=1n(yi-(axi+bi))2 и σ2=∑i=1n(yi-g(xi))2, минимальное значение будет соответствовать более подходящей линии.
σ1=∑i=1n(yi-(axi+bi))2==∑i=15(yi-(0,165xi+2,184))2≈0,019σ2=∑i=1n(yi-g(xi))2==∑i=15(yi-(xi+13+1))2≈0,096
Ответ: поскольку σ1<σ2, то прямой, наилучшим образом аппроксимирующей исходные данные, будет
y=0,165x+2,184.
Нужна помощь преподавателя?
Опиши задание — и наши эксперты тебе помогут!
Описать задание
Как изобразить МНК на графике функций
Метод наименьших квадратов наглядно показан на графической иллюстрации. С помощью красной линии отмечена прямая g(x)=x+13+1, синей – y=0,165x+2,184. Исходные данные обозначены розовыми точками.
Поясним, для чего именно нужны приближения подобного вида.
Они могут быть использованы в задачах, требующих сглаживания данных, а также в тех, где данные надо интерполировать или экстраполировать. Например, в задаче, разобранной выше, можно было бы найти значение наблюдаемой величины y при x=3 или при x=6. Таким примерам мы посвятили отдельную статью.
Доказательство метода МНК
Чтобы функция приняла минимальное значение при вычисленных a и b, нужно, чтобы в данной точке матрица квадратичной формы дифференциала функции вида F(a, b)=∑i=1n(yi-(axi+b))2 была положительно определенной.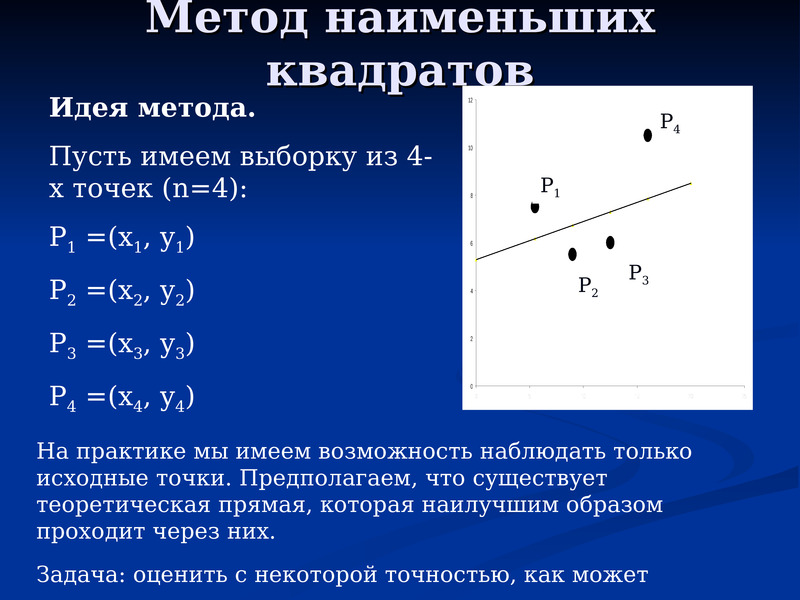 Покажем, как это должно выглядеть.
Покажем, как это должно выглядеть.
Пример 2
У нас есть дифференциал второго порядка следующего вида:
d2F(a; b)=δ2F(a; b)δa2d2a+2δ2F(a; b)δaδbdadb+δ2F(a; b)δb2d2b
Решение
δ2F(a; b)δa2=δδF(a; b)δaδa==δ-2∑i=1n(yi-(axi+b))xiδa=2∑i=1n(xi)2δ2F(a; b)δaδb=δδF(a; b)δaδb==δ-2∑i=1n(yi-(axi+b))xiδb=2∑i=1nxiδ2F(a; b)δb2=δδF(a; b)δbδb=δ-2∑i=1n(yi-(axi+b))δb=2∑i=1n(1)=2n
Иначе говоря, можно записать так: d2F(a; b)=2∑i=1n(xi)2d2a+2·2∑xii=1ndadb+(2n)d2b.
Мы получили матрицу квадратичной формы вида M=2∑i=1n(xi)22∑i=1nxi2∑i=1nxi2n.
В этом случае значения отдельных элементов не будут меняться в зависимости от a и b. Является ли эта матрица положительно определенной? Чтобы ответить на этот вопрос, проверим, являются ли ее угловые миноры положительными.
Вычисляем угловой минор первого порядка: 2∑i=1n(xi)2>0. Поскольку точки xi не совпадают, то неравенство является строгим. Будем иметь это в виду при дальнейших расчетах.
Вычисляем угловой минор второго порядка:
det(M)=2∑i=1n(xi)22∑i=1nxi2∑i=1nxi2n=4n∑i=1n(xi)2-∑i=1nxi2
После этого переходим к доказательству неравенства n∑i=1n(xi)2-∑i=1nxi2>0 с помощью математической индукции.
- Проверим, будет ли данное неравенство справедливым при произвольном n. Возьмем 2 и подсчитаем:
2∑i=12(xi)2-∑i=12xi2=2×12+x22-x1+x22==x12-2x1x2+x22=x1+x22>0
У нас получилось верное равенство (если значения x1 и x2 не будут совпадать).
- Сделаем предположение, что данное неравенство будет верным для n, т.е. n∑i=1n(xi)2-∑i=1nxi2>0 – справедливо.
- Теперь докажем справедливость при n+1, т.е. что (n+1)∑i=1n+1(xi)2-∑i=1n+1xi2>0, если верно n∑i=1n(xi)2-∑i=1nxi2>0.
Вычисляем:
(n+1)∑i=1n+1(xi)2-∑i=1n+1xi2==(n+1)∑i=1n(xi)2+xn+12-∑i=1nxi+xn+12==n∑i=1n(xi)2+n·xn+12+∑i=1n(xi)2+xn+12—∑i=1nxi2+2xn+1∑i=1nxi+xn+12==∑i=1n(xi)2-∑i=1nxi2+n·xn+12-xn+1∑i=1nxi+∑i=1n(xi)2==∑i=1n(xi)2-∑i=1nxi2+xn+12-2xn+1×1+x12++xn+12-2xn+1×2+x22+…+xn+12-2xn+1×1+xn2==n∑i=1n(xi)2-∑i=1nxi2++(xn+1-x1)2+(xn+1-x2)2+…+(xn-1-xn)2>0
Выражение, заключенное в фигурные скобки, будет больше 0 (исходя из того, что мы предполагали в пункте 2), и остальные слагаемые будут больше 0, поскольку все они являются квадратами чисел. Мы доказали неравенство.
Мы доказали неравенство.
Ответ: найденные a и b будут соответствовать наименьшему значению функции F(a, b)=∑i=1n(yi-(axi+b))2, значит, они являются искомыми параметрами метода наименьших квадратов (МНК).
Метод наименьших квадратов (МНК) (Пространственная статистика)—ArcMap
Основным результатом работы этого инструмента является файл отчета, который записывается в окне Результаты. Если щелкнуть правой кнопкой мыши запись Сообщения в окне Результаты и выбрать Вид, итоговый отчет инструмента Исследовательская регрессия можно будет просмотреть в диалоговом окне Сообщение.
Примечание:
Если данный инструмент является частью пользовательского инструмента моделирования, дополнительные таблицы будут отображаться в окне Результаты только в том случае, если перед запуском инструмента они были заданы в качестве параметров модели.
Инструмент МНК также создает выходной класс объектов и дополнительные таблицы с информацией о коэффициентах и диагностике. Все это доступно в окне Результаты. Выходной класс объектов автоматически добавляется в таблицу содержания со схемой отображения горячих/холодных точек, применяемой к невязкам моделей. Полное пояснение по каждому результату см. в разделе Интерпретация результатов по МНК.
Все это доступно в окне Результаты. Выходной класс объектов автоматически добавляется в таблицу содержания со схемой отображения горячих/холодных точек, применяемой к невязкам моделей. Полное пояснение по каждому результату см. в разделе Интерпретация результатов по МНК.
Результаты регрессии МНК являются заслуживающими доверия только в том случае, если ваши данные и модель регрессии удовлетворяет всем допущениям, соответствующим этому методу. Проанализируйте таблицу Распространенные проблемы, последствия и решения регрессии в Основах анализа регрессии, чтобы гарантировать, что ваша модель должным образом определена.
Зависимые и независимые переменные должны храниться в числовых полях, содержащих разнообразие значений. МНК не может работать, когда все переменные имеют одинаковые значения (например, все значения для поля равны 9.0). Линейные методы регрессии, такие, как МНК, не являются подходящими для прогнозирования двоичных результатов (например, все значения для зависимой переменной равны или 1 или 0).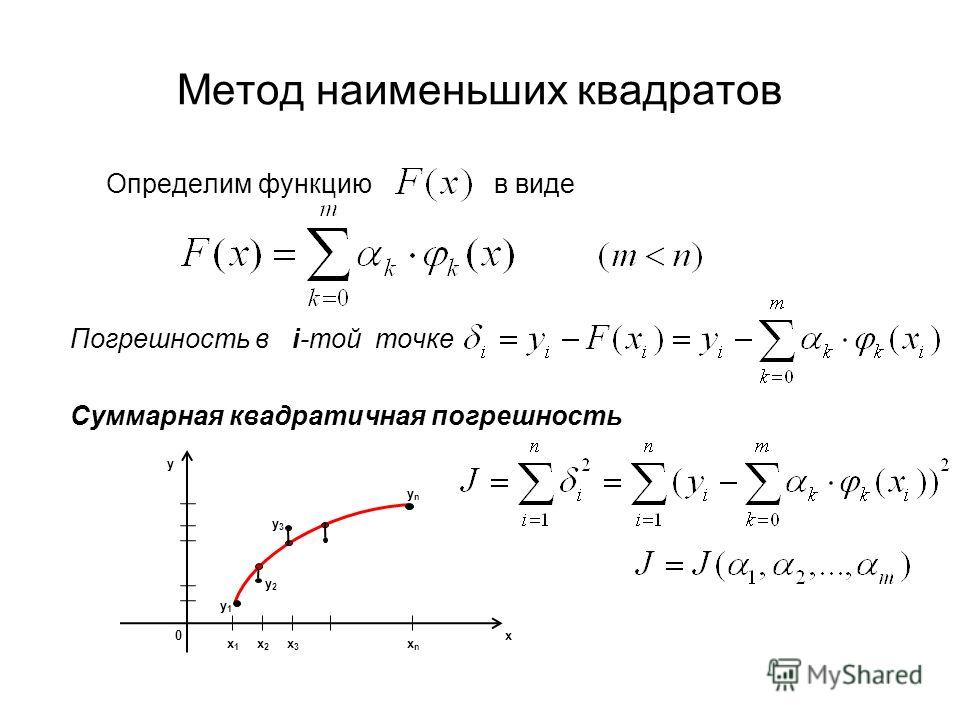
Поле Unique ID связывает прогнозированные значения в модели с каждым объектом. Следовательно, значения поля Unique ID должны быть уникальными для каждого объекта и, как правило, это поле должно быть постоянным полем, принадлежащим классу объектов. Если у вас нет поля Unique ID, вы можете легко создать его путем добавления нового целого поля в вашу таблицу класса объектов и введения значений поля, аналогичных полю FID/OID. Вы не можете использовать поле FID/OID напрямую в параметре Уникальный ID.
Всякий раз, когда есть статистически значимая пространственная автокорреляция невязок регрессии, модель МНК будут считаться неопределенной. Следовательно, результаты регрессии по МНК будут ненадежными. Примените инструмент Пространственная автокорреляция к невязкам вашей регрессии, чтобы оценить потенциальные проблемы. Статистически значимая пространственная автокорреляция невязок регрессии почти всегда указывает на один или несколько недостающих ключевых независимых переменных модели.
Визуально оцените все очевидные смещения прогнозируемых значений в большую и меньшую сторону в невязках вашей регрессии, чтобы увидеть, дают ли они представления о потенциальных недостающих переменных в вашей регрессионной модели. Иногда проведение Анализа горячих точек по невязкам помогает визуализировать пространственную кластеризацию отклонений прогнозируемых значений в большую и меньшую сторону.
Если неопределенность является результатом попытки моделировать нестационарные переменные, используя глобальную модель (МНК – это глобальная модель), то может быть использована Географически взвешенная регрессия для повышения точности прогнозирования и лучшего понимания нестационарности (региональной вариабельности) в ваших независимых переменных.
Если результатом вычисления является бесконечность или неопределенность, результат для файлов, которые не являются шейп-файлами, будет Null; для шейп-файлов результат будет – DBL_MAX (например, -1. 7976931348623158e+308).
7976931348623158e+308).
Итоговые результаты диагностики модели записываются в итоговый отчет по МНК и в дополнительную выходную таблицу результатов диагностики. Обе записи включают результаты диагностики исправленного Информационного критерия Akaike (AICc), коэффициент определения, соединенную F-статистику, статистику Вальда, стьюдентизированную Кенкером статистику Бреуша-Пагана и статистику Жарке-Бера. Диагностическая таблица также включает нескорректированные значения AIC и S-квадрат.
Дополнительный коэффициент и/или диагностические выходные таблицы, если они уже существуют, будут переписаны, если включена опция перезаписи результатов операций геообработки.
Этот инструмент при необходимости создает PDF-файл отчета со сводным представлением результатов. PDF-файл не отображается автоматически в окне Каталога. Чтобы PDF-файлы отображались в окне Каталога, откройте приложение ArcCatalog, выберите опцию меню Настройка, щелкните Опции ArcCatalog и выберите закладку Типы файлов. Нажмите кнопку Новый тип и укажите PDF, как показано ниже, для параметра Расширение файла.
Нажмите кнопку Новый тип и укажите PDF, как показано ниже, для параметра Расширение файла.
На компьютерах с языковыми пакетами ArcGIS для арабского языка и других языков, которые читаются справа налево, в PDF-файле выходного отчета может отсутствовать текст или элементы форматирования. Эти проблемы описаны в этой статье.
Слои карты можно использовать для определения Входного класса объектов. Если в слое есть выборка, только выбранные объекты будут включены в анализ.
Внимание:
При использовании шейп-файлов, помните, что в них нельзя хранить нулевые (null) значения. Инструменты или другие процедуры, создающие шейп-файлы из прочих входных данных, могут хранить значения NULL в виде 0 или оперировать ими как нулем. В некоторых случаях нули в шейп-файлах хранятся как очень маленькие отрицательные числа. Это может привести к неожиданным результатам. Дополнительные сведения см. в разделе Рекомендации по геообработке выходных данных шейп-файла.
в разделе Рекомендации по геообработке выходных данных шейп-файла.
|
Экстраполяция — это метод научного исследования, который основан на распространении прошлых
Сущность метода наименьших квадратов состоит в минимизации суммы квадратических отклонений между
Теоретический анализ сущности изучаемого явления, изменение которого отображается временным рядом, служит основой для выбора
Рабочая формула метода наименьших квадратов: У t+1 = а*Х + b, где t + 1 – прогнозный
Расчет коэффициентов a и b осуществляется по следующим формулам:
где, Уф – фактические значения ряда динамики; n – число уровней временного ряда;
Сглаживание временных рядов методом наименьших квадратов служит для отражения закономерности развития изучаемого явления.
Развитие явления зависит не от того, сколько лет прошло с отправного момента, а от того, какие факторы влияли на его
Правильно установить тип кривой, тип аналитической зависимости от времени – одна из самых сложных задач
Подбор вида функции, описывающей тренд, параметры которой определяются методом наименьших квадратов, производится в
где Уф – фактические значения ряда динамики; Ур – расчетные (сглаженные) значения ряда динамики; n – число уровней временного
Недостатки метода наименьших квадратов:
Пример применения метода наименьших квадратов для разработки прогноза
Задача. Имеются данные, характеризующие уровень безработицы в регионе, %
Решение методом наименьших квадратов
Для решения составим таблицу, в которой будем производить необходимые расчеты:
Определим условное обозначение времени как последовательную нумерацию периодов базы прогноза (графа 3).
Коэффициенты a и b определим по следующим формулам:
где, Уф – фактические значения ряда динамики; n – число уровней временного ряда.
Далее определяем прогнозное значение:
Рассчитываем среднюю относительную ошибку по формуле:
ε = 28,63/10 = 2,86% точность прогноза высокая.
Вывод: Сравнивая результаты, полученные при расчетах
В первом и третьем случае точность прогноза является высокой, поскольку средняя относительная
Другие статьи по данной теме:
Список использованных источников
|







Метод наименьших квадратов, МНК, описание метода наименьших квадратов
В большинстве экспериментальных данных, задаваемых с помощью табличной функции, имеется достаточно большой разброс точек. При этом использование кусочной или непрерывной интерполяции не всегда оправдано, поскольку ставится задача исследовать общую тенденцию изменения физической величины.
В этом общем случае аппроксимации данных искомая кривая не обязательно должна проходить через заданные точки.
Рассмотрим рис. 1, отражающий большой разброс точек. В простейшем случае будем искать аппроксимирующую функцию ф(х) в виде полинома первой степени (прямой):
Рис. 1. Аппроксимация
Таким образом, данная система точек группируется вокруг искомой прямой. Эту прямую легко провести на глаз так, чтобы она наиболее близко подходила к исходным точкам. Однако можно найти уравнение прямой более строгими математическими методами.
Однако можно найти уравнение прямой более строгими математическими методами.
Метод наименьших квадратов наиболе часто используют для решения контрольных по эконометрике для нахождения параметров уравнений (линий, степенной функции, гиперболы и т.д.)
Пусть общее количество точек равно n.
Отклонение i-й точки от искомой прямой:
Как видно из рис. 2, отклонения могут быть как положительными, так и отрицательными. Поэтому для того, чтобы определить близость искомой функции к табличным точкам, необходимо составить сумму квадратов всех отклонений.
Метод наименьших квадратов заключается в минимизации суммы квадратов отклонений. В нашем случае эта функция равна
Рис. 2. График отклонения
Для нахождения минимума функции S необходимо приравнять нулю ее частные производные. В результате получим систему уравнений:
Опуская промежуточные преобразования, получим систему уравнений для нахождения неизвестных коэффициентов:
Здесь m — количество точек; суммирование здесь и далее предполагается по всем точкам.
Метод наименьших квадратов несложно распространить на общий случай, когда мы будем искать функцию ф(х) в виде полинома степени n:
Отметим, что в случае аппроксимации всегда справедливо следующее соотношение, связывающее количество исходных точек m и степень искомого полинома:
причем в случае равенства мы приходим к интерполяции (все отклонения равны нулю).
Неизвестные коэффициенты а находим из условия минимизации суммы квадратов отклонений искомой функции от исходных точек. По аналогии с полиномом первой степени в нашем случае имеем систему уравнений: Z*A = B
где Z — квадратная матрица размерностью (n+1)х(n+1), составленная из известных координат точек, А — вектор неизвестных коэффициентов; Y- вектор-столбец свободных членов.
Применение метода наименьших квадратов в Excel: пример
Программа Excel – мощный табличный редактор, позволяющий выполнять огромное количество различных операций и задач. В данной статье мы разберем, как можно применить метод наименьших квадратов (МНК), который используется для решения различных задач с минимизацией суммы квадратов отклонений некоторых функций от искомых переменных.
В данной статье мы разберем, как можно применить метод наименьших квадратов (МНК), который используется для решения различных задач с минимизацией суммы квадратов отклонений некоторых функций от искомых переменных.
Подготовительный этап: активируем надстройку “Поиск Решения”
Прежде, чем приступить к решению основной задачи, потребуется активировать надстройку “Поиск решения” в программе.
- Идем в меню “Файл”.
- В перечне слева выбираем пункт “Параметры”.
- В правой части подраздела “Надстройки” выбираем для параметра “Управление” вариант “Надстройки Excel” и жмем “Перейти”.
- Появится окно для выбора нужных надстроек. Устанавливаем галочку напротив пункта “Поиск решения” и щелкаем OK.
Этап 1: исходные данные
Давайте разберем применение метода наименьших квадратов, решив конкретный пример. Допустим, у нас есть два ряда числовых значений – X и Y.
Данная зависимость может быть описана уравнением ниже:
Y=A+NX
Также, мы знаем, что если X=0, то и Y=0. А значит, данное уравнение можно записать так:
Y=NX
Приступим к выполнению нашей задачи, которая заключается в нахождении суммы квадратов разности.
Этап 2: решаем задачу с применением МНК
- Столбцу, находящемся слева от X, задаем имя N пишем число “1” (примерное значение первого коэф. N) напротив первого значения ряда X.
- Столбцу с правой стороны от Y задаем название NX. Затем в самой верхней ячейке (напротив первых значений рядов X и Y) пишем формулу произведения коэф. N на соответствующее ему значение из столбца X. При этом адрес ячейки с коэффициентом нужно сделать абсолютным, чтобы он не менялся при копировании формулы. По готовности жмем Enter.
- Наводим указатель мыши на ячейку с полученным результатом.
 Как только появится черный плюсик (маркер заполнения), зажав левую кнопку мыши тянем его вниз до последней строки таблицы.
Как только появится черный плюсик (маркер заполнения), зажав левую кнопку мыши тянем его вниз до последней строки таблицы. - Получаем результаты расчетов в каждой ячейке столбца NX.
- Теперь нужно посчитать сумму разностей квадратов значений Y и NX. Встаем в самую верхнюю ячейку столбца справа от NX (не считая шапки таблицы) и щелкаем по значку “Вставить функцию” (fx).
- В окне вставки функции выбираем категорию “Математические”, находим оператор “СУММКВРАЗН” и щелкаем OK.
- Теперь нужно заполнить аргументы функции:
- в поле “Массив_x” указываем координаты диапазона ячеек столбца Y (без шапки). Адреса ячеек можно указать как вручную, напечатав их с клавиатуры, так и путем выделения с помощью зажатой левой кнопки мыши в самой таблице.
- в поле “Массив_y” указываем диапазон ячеек столбца NX.
- жмем Enter, когда все готово.
- Переключаемся во вкладку “Данные”. В группе “Анализ” щелкаем по функции “Поиск решения”.
- Нам предстоит заполнить параметры поиска решения:
- в поле “Оптимизировать целевую функцию” следует указать ссылку на ячейку с функцией “СУММКВРАЗН”. Сделать это можно вручную или выбрав элемент в таблице.
- для опции “До” выбираем вариант – “Минимум”.
- в поле “Изменяя ячейки переменных” нужно указать координаты ячейки, в которой находится соответствующее значение коэф. N.
- по готовности нажимаем “Найти решение”.
- После выполнения функции появится окно с результатами поиска решения и произойдет замена значения в столбце N. Найденная величина является наименьшим квадратом функции. Нажимаем OK, если полученный результат удовлетворителен.
Заключение
Итак, мы только что разобрали на практическом примере, каким образом можно применить метод наименьших квадратов в Эксель. На практике могут встречаться более сложные задачи, однако, в целом логика действий схожа с той, что мы описали.
На практике могут встречаться более сложные задачи, однако, в целом логика действий схожа с той, что мы описали.
Метод наименьших квадратов.
Поможем написать любую работу на аналогичную
тему
Получить выполненную работу или консультацию специалиста по вашему
учебному проекту
Узнать стоимость
Сущность метода наименьших квадратов заключается в отыскании параметров модели тренда, которая лучше всего описывает тенденцию развития какого-либо случайного явления во времени или в пространстве (тренд – это линия, которая и характеризует тенденцию этого развития). Задача метода наименьших квадратов (МНК) сводится к нахождению не просто какой-то модели тренда, а к нахождению лучшей или оптимальной модели. Эта модель будет оптимальной, если сумма квадратических отклонений между наблюдаемыми фактическими величинами и соответствующими им расчетными величинами тренда будет минимальной (наименьшей):
(9. 1)
1)
где — квадратичное отклонение между наблюдаемой фактической величиной
и соответствующей ей расчетной величиной тренда,
— фактическое (наблюдаемое) значение изучаемого явления,
— расчетное значение модели тренда,
— число наблюдений за изучаемым явлением.
МНК самостоятельно применяется довольно редко. Как правило, чаще всего его используют лишь в качестве необходимого технического приема при корреляционных исследованиях. Следует помнить, что информационной основой МНК может быть только достоверный статистический ряд, причем число наблюдений не должно быть меньше 4-х, иначе, сглаживающие процедуры МНК могут потерять здравый смысл.
Инструментарий МНК сводится к следующим процедурам:
Первая процедура. Выясняется, существует ли вообще какая-либо тенденция изменения результативного признака при изменении выбранного фактора-аргумента, или другими словами, есть ли связь между «у» и «х».
Вторая процедура. Определяется, какая линия (траектория) способна лучше всего описать или охарактеризовать эту тенденцию.
Третья процедура. Рассчитываются параметры регрессионного уравнения, характеризующего данную линию, или другими словами, определяется аналитическая формула, описывающая лучшую модель тренда.
Пример. Допустим, мы имеем информацию о средней урожайности подсолнечника по исследуемому хозяйству (табл. 9.1).
Таблица 9.1
Номер наблюдения | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
Годы | 1995 | 1996 | 1997 | 1998 | 1999 | 2000 | 2001 | 2002 | 2003 | 2004 |
Урожайность, ц/га | 14,2 | 15,6 | 17,5 | 14,5 | 15,3 | 17,0 | 16,6 | 17,5 | 15,0 | 17,7 |
Поскольку уровень технологии при производстве подсолнечника в нашей стране за последние 10 лет практически не изменился, значит, по всей видимости, колебания урожайности в анализируемый период очень сильно зависели от колебания погодно-климатических условий. Действительно ли это так?
Действительно ли это так?
Первая процедура МНК. Проверяется гипотеза о существовании тенденции изменения урожайности подсолнечника в зависимости от изменения погодно-климатических условий за анализируемые 10 лет.
В данном примере за «y» целесообразно принять урожайность подсолнечника, а за «x» – номер наблюдаемого года в анализируемом периоде. Проверку гипотезы о существовании какой-либо взаимосвязи между «x» и «y» можно выполнить двумя способами: вручную и при помощи компьютерных программ. Конечно, при наличии компьютерной техники данная проблема решается сама собой. Но, чтобы лучше понять инструментарий МНК целесообразно выполнить проверку гипотезы о существовании связи между «x» и «y» вручную, когда под рукой находятся только ручка и обыкновенный калькулятор. В таких случаях гипотезу о существовании тенденции лучше всего проверить визуальным способом по расположению графического изображения анализируемого ряда динамики — корреляционного поля:
Корреляционное поле в нашем примере расположено вокруг медленно возрастающей линии. Это уже само по себе говорит о существовании определенной тенденции в изменении урожайности подсолнечника. Нельзя говорить о наличии какой-либо тенденции лишь тогда, когда корреляционное поле похоже на круг, окружность, строго вертикальное или строго горизонтальное облако, или же состоит из хаотично разбросанных точек. Во всех остальных случаях следует подтвердить гипотезу о существовании взаимосвязи между «x» и «y», и продолжить исследования.
Это уже само по себе говорит о существовании определенной тенденции в изменении урожайности подсолнечника. Нельзя говорить о наличии какой-либо тенденции лишь тогда, когда корреляционное поле похоже на круг, окружность, строго вертикальное или строго горизонтальное облако, или же состоит из хаотично разбросанных точек. Во всех остальных случаях следует подтвердить гипотезу о существовании взаимосвязи между «x» и «y», и продолжить исследования.
Вторая процедура МНК. Определяется, какая линия (траектория) способна лучше всего описать или охарактеризовать тенденцию изменения урожайности подсолнечника за анализируемый период.
При наличии компьютерной техники подбор оптимального тренда происходит автоматически. При «ручной» обработке выбор оптимальной функции осуществляется, как правило, визуальным способом – по расположению корреляционного поля. То есть, по виду графика подбирается уравнение линии, которая лучше всего подходит к эмпирическому тренду (к фактической траектории).
Как известно, в природе существует огромное разнообразие функциональных зависимостей, поэтому визуальным способом проанализировать даже незначительную их часть — крайне затруднительно. К счастью, в реальной экономической практике большинство взаимосвязей достаточно точно могут быть описаны или параболой, или гиперболой, или же прямой линией. В связи с этим, при «ручном» варианте подбора лучшей функции, можно ограничиться только этими тремя моделями.
Прямая: |
| Гипербола: |
|
Парабола второго порядка: :
Нетрудно заметить, что в нашем примере лучше всего тенденцию изменения урожайности подсолнечника за анализируемые 10 лет характеризует прямая линия, поэтому уравнением регрессии будет уравнение прямой.
Третья процедура. Рассчитываются параметры регрессионного уравнения, характеризующего данную линию, или другими словами, определяется аналитическая формула, описывающая лучшую модель тренда.
Рассчитываются параметры регрессионного уравнения, характеризующего данную линию, или другими словами, определяется аналитическая формула, описывающая лучшую модель тренда.
Нахождение значений параметров уравнения регрессии, в нашем случае параметров и , является сердцевиной МНК. Данный процесс сводится к решению системы нормальных уравнений.
(9.2)
Эта система уравнений довольно легко решается методом Гаусса. Напомним, что в результате решения, в нашем примере, находятся значения параметров и . Таким образом, найденное уравнение регрессии будет иметь следующий вид:
В линейном уравнении параметр – коэффициент регрессии указывает, на сколько единиц в среднем изменится с изменением на единицу. Он имеет единицу измерения результативного признака. В случае прямой связи – величина положительная, а при обратном – отрицательная. Параметр – свободный член уравнения регрессии, то есть это значениепри . Если не получает нулевых значений, этот параметр имеет лишь расчетное назначение.
Если не получает нулевых значений, этот параметр имеет лишь расчетное назначение.
Приведем также системы нормальных уравнений для отыскивания параметров нелинейных уравнений.
Таблица 9.2
Форма связи | Уравнение связи | Система нормальных уравнений |
параболическая | ||
гиперболическая |
Следует помнить, что при изменении хотя бы одного значения входных данных (пары значенийили одного из них) все коэффициенты изменят в общем случае свои значения, потому что они полностью определяются входными данными. Поэтому при повторной аппроксимации с несколькими измененными данными будет получена другая аппроксимирующая функция с другими коэффициентами.
Внимание!
Если вам нужна помощь в написании работы, то рекомендуем обратиться к
профессионалам. Более 70 000 авторов готовы помочь вам прямо сейчас. Бесплатные
корректировки и доработки. Узнайте стоимость своей работы.
примеры и вычисление функции потерь
Линейная регрессия (Linear regression) — модель зависимости переменной x от одной или нескольких других переменных (факторов, регрессоров, независимых переменных) с линейной функцией зависимости.
Линейная регрессия относится к задаче определения «линии наилучшего соответствия» через набор точек данных и стала простым предшественником нелинейных методов, которые используют для обучения нейронных сетей. В этой статье покажем вам примеры линейной регрессии.
Применение линейной регрессии
Предположим, нам задан набор из 7 точек (таблица ниже).
Цель линейной регрессии — поиск линии, которая наилучшим образом соответствует этим точкам. Напомним, что общее уравнение для прямой есть f (x) = m⋅x + b, где m — наклон линии, а b — его y-сдвиг. Таким образом, решение линейной регрессии определяет значения для m и b, так что f (x) приближается как можно ближе к y. Попробуем несколько случайных кандидатов:
Довольно очевидно, что первые две линии не соответствуют нашим данным. Третья, похоже, лучше, чем две другие. Но как мы можем это проверить? Формально нам нужно выразить, насколько хорошо подходит линия, и мы можем это сделать, определив функцию потерь.
Функция потерь — метод наименьших квадратов
Функция потерь — это мера количества ошибок, которые наша линейная регрессия делает на наборе данных. Хотя есть разные функции потерь, все они вычисляют расстояние между предсказанным значением y(х) и его фактическим значением. Например, взяв строку из среднего примера выше, f(x)=−0.11⋅x+2.5, мы выделяем дистанцию ошибки между фактическими и прогнозируемыми значениями красными пунктирными линиями.
Например, взяв строку из среднего примера выше, f(x)=−0.11⋅x+2.5, мы выделяем дистанцию ошибки между фактическими и прогнозируемыми значениями красными пунктирными линиями.
Одна очень распространенная функция потерь называется средней квадратичной ошибкой (MSE). Чтобы вычислить MSE, мы просто берем все значения ошибок, считаем их квадраты длин и усредняем.
Вычислим MSE для каждой из трех функций выше: первая функция дает MSE 0,17, вторая — 0,08, а третья — 0,02. Неудивительно, что третья функция имеет самую низкую MSE, подтверждая нашу догадку, что это линия наилучшего соответствия.
Рассмотрим приведенный ниже рисунок, который использует две визуализации средней квадратичной ошибки в диапазоне, где наклон m находится между -2 и 4, а b между -6 и 8.
Слева: диаграмма, изображающая среднеквадратичную ошибку для -2≤m≤4, -6≤p≤8 Справа: тот же рисунок, но визуализирован как контурный график, где контурные линии являются логарифмически распределенными поперечными сечениями высоты.
Глядя на два графика, мы видим, что наш MSE имеет форму удлиненной чаши, которая, по-видимому, сглаживается в овале, грубо центрированном по окрестности (m, p) ≈ (0.5, 1.0). Если мы построим MSE линейной регрессии для другого датасета, то получим аналогичную форму. Поскольку мы пытаемся минимизировать MSE, наша цель — выяснить, где находится самая низкая точка в чаше.
Больше размерностей
Вышеприведенный пример очень простой, он имеет только одну независимую переменную x и два параметра m и b. Что происходит, когда имеется больше переменных? В общем случае, если есть n переменных, их линейная функция может быть записана как:
f(x) = b+w_1*x_1 + … + w_n*x_n
Один трюк, который применяют, чтобы упростить это — думать о нашем смещении «b», как о еще одном весе, который всегда умножается на «фиктивное» входное значение 1. Другими словами:
f(x) = b*1+w_1*x_1 + … + w_n*x_n
Добавление измерений, на первый взгляд, ужасное усложнение проблемы, но оказывается, постановка задачи остается в точности одинаковой в 2, 3 или в любом количестве измерений. Существует функция потерь, которая выглядит как чаша — гипер-чаша! И, как и прежде, наша цель — найти самую нижнюю часть этой чаши, объективно наименьшее значение, которое функция потерь может иметь в отношении выбора параметров и набора данных.
Существует функция потерь, которая выглядит как чаша — гипер-чаша! И, как и прежде, наша цель — найти самую нижнюю часть этой чаши, объективно наименьшее значение, которое функция потерь может иметь в отношении выбора параметров и набора данных.
Итак, как мы вычисляем, где именно эта точка на дне? Распространенный подход — обычный метод наименьших квадратов, который решает его аналитически. Когда есть только один или два параметра для решения, это может быть сделано вручную, и его обычно преподают во вводном курсе по статистике или линейной алгебре.
Проклятие нелинейности
Увы, обычный МНК не используют для оптимизации нейронных сетей, поэтому решение линейной регрессии будет оставлено как упражнение, оставленное читателю. Причина, по которой линейную регрессию не используют, заключается в том, что нейронные сети нелинейны.
Различие между линейными уравнениями, которые мы составили, и нейронной сетью — функция активации (например, сигмоида, tanh, ReLU или других).
Эта нелинейность означает, что параметры не действуют независимо друг от друга, влияя на форму функции потерь. Вместо того, чтобы иметь форму чаши, функция потерь нейронной сети более сложна. Она ухабиста и полна холмов и впадин. Свойство быть «чашеобразной» называется выпуклостью, и это ценное свойство в многопараметрической оптимизации. Выпуклая функция потерь гарантирует, что у нас есть глобальный минимум (нижняя часть чаши), и что все дороги под гору ведут к нему.
Минимум функции
Но, вводя нелинейность, мы теряем это удобство ради того, чтобы дать нейронным сетям гораздо большую «гибкость» при моделировании произвольных функций. Цена, которую мы платим, заключается в том, что больше нет простого способа найти минимум за один шаг аналитически. В этом случае мы вынуждены использовать многошаговый численный метод, чтобы прийти к решению. Хотя существует несколько альтернативных подходов, градиентный спуск остается самым популярным методом.
mnk-1 — Перевод на английский — примеры французский
Французский
арабский
Немецкий
английский
испанский
Французский
иврит
Итальянский
Японский
нидерландский язык
Польский
португальский
румынский
русский
турецкий
китайский язык
английский
Синонимы
арабский
Немецкий
английский
испанский
Французский
иврит
Итальянский
Японский
нидерландский язык
Польский
португальский
румынский
русский
турецкий
китайский язык
Эти примеры могут содержать грубые слова, основанные на вашем поиске.
Эти примеры могут содержать разговорные слова, основанные на вашем поиске.
Другие переводы
СТРУКТУРА КРИСТАЛЛОГРАФИЧЕСКИХ БЕЛКОВ MNK-1 ET MNK-2
КРИСТАЛЛОГРАФИЧЕСКАЯ СТРУКТУРА БЕЛКОВ МНК-1 И МНК-2
Изобретение касается кристаллических киназ Mnk-1, и Mnk-2 и др., А также уточняется, что кристаллическая структура доменных киназ Mnk-1, и Mnk-2.
Настоящее изобретение относится к кристаллическим киназам Mnk-1, и Mnk-2 и, в частности, к кристаллической структуре киназного домена Mnk-1 и Mnk-2.
Предложите пример
Другие результаты
структура cristallographique des protéines mnk — 1 et mnk -2
кристаллографическая структура белков mnk — 1 и mnk -2
Нет результатов для этого значения. Показать больше примеров
Показать больше примеров
Результатов: 3. Точное: 2. Затраченное время: 129 мс.
Что нужно знать: SiegeAcademy
* Заявление об ограничении ответственности * Я ни в коем случае не выступаю за использование MnK на консолях. Это дает вам вопиющее, несправедливое преимущество, и в целом это просто отстой. Кроме того, у меня мало опыта против MnK на более низких уровнях, поэтому я понятия не имею, каково это там.
Я сам играю на консолях (Xbox) и видел определенный рост количества игроков, использующих MnK на более высоком уровне (более высокий уровень и алмаз).Из 3-4 игр, которые моя команда обычно играет за одну сессию, мы сталкиваемся как минимум с 1 человеком, использующим мышь, и, по крайней мере, каждые 10 или около того игр мы сталкиваемся с пятью стеками MnK. В связи с появлением большого количества сообщений в основной подпункте и этой о MnK, я хочу поделиться некоторыми вещами, которые я узнал:
MnK-плееров не так много, как вы могли бы подумать.
 Хотя я заметил увеличение количества людей, использующих MnK, это также связано с тем, что я стал лучше в игре и играю против людей с более высоким рейтингом MMR.Если вы сидите на уровне Gold или ниже, вы не столкнетесь с почти таким же количеством пользователей MnK, как тот, кто находится на уровне Plat 1 или Diamond. Там ваша главная проблема — тролли и люди, играющие на убийства, а не на цель. Кроме того, не спешите говорить, что у кого-то есть мышь. Требуется немного опыта в игре, чтобы действительно точно определить движение MnK по сравнению с высокой чувствительностью. Я играю на обычном контроллере, и меня обвиняли в том, что я играю на MnK, хотя я признаю, что моя механика НЕ привела меня к алмазу.Иногда это очевидно, но во многих других случаях это сложно сказать.
Хотя я заметил увеличение количества людей, использующих MnK, это также связано с тем, что я стал лучше в игре и играю против людей с более высоким рейтингом MMR.Если вы сидите на уровне Gold или ниже, вы не столкнетесь с почти таким же количеством пользователей MnK, как тот, кто находится на уровне Plat 1 или Diamond. Там ваша главная проблема — тролли и люди, играющие на убийства, а не на цель. Кроме того, не спешите говорить, что у кого-то есть мышь. Требуется немного опыта в игре, чтобы действительно точно определить движение MnK по сравнению с высокой чувствительностью. Я играю на обычном контроллере, и меня обвиняли в том, что я играю на MnK, хотя я признаю, что моя механика НЕ привела меня к алмазу.Иногда это очевидно, но во многих других случаях это сложно сказать.Игроки MnK не обязательно плохо играют. Признайтесь, если вы картофель на палочках, покупка мыши не сделает вас богом. Большинство игроков MnK обладают наполовину приличным чувством игры и, вероятно, находятся на уровне, близком или выше Plat 3 на контроллере.
 Хотя MnK определенно помогли им продвинуться дальше, они не мусор в игре. Это мерзко? Да. Были бы они так же хороши на ПК? Может быть.Они обманывают? Технически, да (против правил ESL, и я думаю, что Ubi сказал, что это так). Вы бы победили их, если бы у них был контроллер? Опять же, может быть. Дело в том, что они, скорее всего, не поднялись с серебра на плато / бриллиант только за счет MnK. Помните, они дерьмовые люди, а не дерьмовые игроки.
Хотя MnK определенно помогли им продвинуться дальше, они не мусор в игре. Это мерзко? Да. Были бы они так же хороши на ПК? Может быть.Они обманывают? Технически, да (против правил ESL, и я думаю, что Ubi сказал, что это так). Вы бы победили их, если бы у них был контроллер? Опять же, может быть. Дело в том, что они, скорее всего, не поднялись с серебра на плато / бриллиант только за счет MnK. Помните, они дерьмовые люди, а не дерьмовые игроки.Чтобы оттолкнуться от второй точки, многие игроки MnK занимаются аимбро. Они с большей вероятностью будут играть агрессивно, делать то, что не делает нормальный человек, и продвигаться быстрее, чем обычная команда.Это может быть утомительно, но готовность к каждому агрессивному пику или игре очень поможет вам, и это выходит за рамки игроков MnK.
Это также относится к случаям, когда люди целятся лучше, чем вы в целом, но против MnK чрезмерно агрессивная игра вам не подходит. Играть нужно максимально умно и консервативно.
 Не участвуйте в перестрелках, которые вам не нужны. Если вы находитесь в роуминге и испытываете давление, обратитесь за помощью или уходите оттуда как можно скорее. Не бросай свою жизнь.Держите углы и старайтесь быть непредсказуемым. В обороне или на постпроизводстве время — ваш друг, и вы должны его использовать. Когда вы атакуете, работайте как можно лучше в команде и играйте на цель, а не на убийства. Вы должны стремиться к чистым играм и хорошей командной работе, и игра против MnK — действительно хорошая практика для этого.
Не участвуйте в перестрелках, которые вам не нужны. Если вы находитесь в роуминге и испытываете давление, обратитесь за помощью или уходите оттуда как можно скорее. Не бросай свою жизнь.Держите углы и старайтесь быть непредсказуемым. В обороне или на постпроизводстве время — ваш друг, и вы должны его использовать. Когда вы атакуете, работайте как можно лучше в команде и играйте на цель, а не на убийства. Вы должны стремиться к чистым играм и хорошей командной работе, и игра против MnK — действительно хорошая практика для этого.Скорее всего, вы умерли не потому, что у них была мышь. Убедитесь, что вы, когда вас убьют, думаете о том, что вы могли бы сделать лучше в этой ситуации.Иногда вас просто переигрывали, и вы в любом случае умерли бы. Конечно, бывают случаи, когда вы проигрывали в перестрелке или погибали просто потому, что у них была мышь, но если вы постоянно обвиняете мышь, вам будет легче наклоняться.
По возможности не играйте поздно ночью.
 Из-за некоторых вещей в расписании моя команда не может играть до 7 или 8, а мы играем примерно до 11 или 12. Это, вероятно, худшее время для игры, если вы хотите избежать MnK, поскольку обычно игры становятся более потными, чем позже вы начинаете. играет.Я знаю, что заметил значительно больше MnK ночью, чем днем, поэтому, если вы можете играть днем, у вас может быть меньше проблем. Может быть, это только для меня, но может помочь другим.
Из-за некоторых вещей в расписании моя команда не может играть до 7 или 8, а мы играем примерно до 11 или 12. Это, вероятно, худшее время для игры, если вы хотите избежать MnK, поскольку обычно игры становятся более потными, чем позже вы начинаете. играет.Я знаю, что заметил значительно больше MnK ночью, чем днем, поэтому, если вы можете играть днем, у вас может быть меньше проблем. Может быть, это только для меня, но может помочь другим.
Игроки MnK не непобедимы, и на консолях это легко. Нам не нужно иметь дело с хакерами, и с партийной системой очень просто избежать любого вида чата. Играйте с умом, не бросайте свою жизнь попусту и играйте в команде, и вам будет намного легче победить MnK.
(Не стесняйтесь искать меня на R6Tab.Мой главный герой — Рака Сируси. У меня есть команда, поэтому я не собираюсь объединяться.)
Примеры Ace [BRD, MNK] [Требуется аддон]
Профили, которые я активно использую со своим аддоном. Считайте их примерами, если вам нужны ссылки / базовые нагрузки, которые можно использовать для настройки, если вы используете мой аддон. Они не будут работать без моего дополнения (они много работают, но некоторые тонкости будут потеряны).
Они не будут работать без моего дополнения (они много работают, но некоторые тонкости будут потеряны).
Не стесняйтесь задавать вопросы, спасибо.
Спасибо, ас 🙂 Чем больше профилей мы здесь получим, тем лучше! Как и требовал автор аддона, можно только предполагать, что это вершина крутизны>.>
профиль монаха ДЕЙСТВИТЕЛЬНО хороший, держит меня на одном уровне или чуть ниже некоторых очень хороших drgs … мне не хватает ключевого снаряжения, поэтому я могу даже быть выше них oO
Я могу быть идиотом, но есть ли аддон под названием ace’s addon?
[Требуется аддон] не уверен, о каком аддоне идет речь.
это старый взгляд на марш человека марша потока. с тех пор появилась полностью новая версия бота. аддона ace больше нет, потому что многие из этих функций уже есть в боте.
да, я так много думал, я знал, что он старый. Спасибо за ответ Джеки, ты был только полезным ….;)
, они могут даже работать из-за изменений, добавленных в диспетчер навыков в большом обновлении, без вреда в их удалении и испытании.
вы понимаете, что это действительно очень старый? Ace тем временем делает включенные профили, я думаю, они хорошо работают
(12.02.2015, 00:41) roberto_mm86 Написал: [->] Heyla!
Ty ace, поделитесь своими 2 профилями!
Я попробовал профиль BRD и работает ДЕЙСТВИТЕЛЬНО хорошо !!!
Но у меня всего 1 вопрос.
Фильтры = горячие клавиши для разных вращений.
Например, Фильтр 1 Вращение одной цели. Фильтр 2 Вращение по площади.
, поэтому в режиме помощи вы можете отключить фильтр 2 для борьбы с боссами, а когда вам нужно Aoe, вы можете отключить фильтр 1 и включить фильтр 2. они устанавливаются с помощью комбинаций горячих клавиш, которые вы делаете на клавиатуре.
Перевести mnk на французский язык с контекстными примерами
Вклад человека
От профессиональных переводчиков, предприятий, веб-страниц и бесплатных хранилищ переводов.
Добавить перевод
Французский
Малади де Менкес
Последнее обновление: 2014-12-09
Частота использования: 1
Качество:
Предупреждение: Это выравнивание может быть неправильным.
Удалите, пожалуйста, вы так считаете.
Французский
Синдром де Менкеса
Последнее обновление: 2014-12-09
Частота использования: 5
Качество:
Предупреждение: Это выравнивание может быть неправильным.
Удалите, пожалуйста, вы так считаете.
Французский
Синдром де Менкеса
Последнее обновление: 2014-12-09
Частота использования: 1
Качество:
Предупреждение: Это выравнивание может быть неправильным.
Удалите, пожалуйста, вы так считаете.
Французский
Синдром des cheveux crépus
Последнее обновление: 2014-12-09
Частота использования: 1
Качество:
Предупреждение: Это выравнивание может быть неправильным.
Удалите, пожалуйста, вы так считаете.
Французский
Синдром des cheveux en fil de fer
Последнее обновление: 2014-12-09
Частота использования: 1
Качество:
Предупреждение: Это выравнивание может быть неправильным.
Удалите, пожалуйста, вы так считаете.
Последнее обновление: 2011-07-27
Частота использования: 1
Качество:
Последнее обновление: 2018-02-13
Частота использования: 1
Качество:
Английский
МНК — синдром Менкеса
Французский
Малади де Менкес
Последнее обновление: 2014-12-09
Частота использования: 2
Качество:
Предупреждение: Это выравнивание может быть неправильным.
Удалите, пожалуйста, вы так считаете.
Английский
МНК — синдром Менкеса
Французский
Синдром де Менкеса
Последнее обновление: 2014-12-09
Частота использования: 10
Качество:
Предупреждение: Это выравнивание может быть неправильным.
Удалите, пожалуйста, вы так считаете.
Английский
МНК — синдром Менкеса
Французский
Синдром де Менкеса
Последнее обновление: 2014-12-09
Частота использования: 2
Качество:
Предупреждение: Это выравнивание может быть неправильным.
Удалите, пожалуйста, вы так считаете.
Английский
МНК — синдром Менкеса
Французский
Синдром des cheveux crépus
Последнее обновление: 2014-12-09
Частота использования: 2
Качество:
Предупреждение: Это выравнивание может быть неправильным.
Удалите, пожалуйста, вы так считаете.
Английский
МНК — синдром Менкеса
Французский
Синдром des cheveux en fil de fer
Последнее обновление: 2014-12-09
Частота использования: 2
Качество:
Предупреждение: Это выравнивание может быть неправильным.
Удалите, пожалуйста, вы так считаете.
Английский
лечение рака путем модуляции мнк
Французский
traitement du Cancer par modulation d’un mnk
Последнее обновление: 2011-07-27
Частота использования: 1
Качество:
Английский
ЛЕЧЕНИЕ РАКА С ПОМОЩЬЮ МОДУЛЯЦИИ MNK
Французский
ТРАКТЕРИСТИКА РАКА PAR МОДУЛЯЦИЯ D’UN MNK
Последнее обновление: 2011-07-27
Частота использования: 1
Качество:
Английский
ПИРИДИН И ПРОИЗВОДНЫЕ ПИРАЗИНА КАК ИНГИБИТОРЫ MNK-КИНАЗ
Французский
DÉRIVÉS DE PYRIDINE ET DE PYRAZINE UTILISÉS EN TANT QU & # 39; INHIBITEURS DE LA KINASE MNK
Последнее обновление: 2014-11-25
Частота использования: 4
Качество:
Предупреждение: содержит невидимое форматирование HTML
Английский
производные пиридина и пиразина как ингибиторы мнк-киназы
Французский
производные пиридина и пиразина, используемые в качестве ингибиторов киназы mnk
Последнее обновление: 2011-07-27
Частота использования: 1
Качество:
Английский
КРИСТАЛЛОГРАФИЧЕСКАЯ СТРУКТУРА БЕЛКОВ МНК-1 И МНК-2
Французский
СТРУКТУРА КРИСТАЛЛОГРАФИЧЕСКИХ БЕЛКОВ MNK-1 ET MNK-2
Последнее обновление: 2014-11-25
Частота использования: 11
Качество:
Английский
использование ингибиторов mnk для лечения болезни Альцгеймера
Французский
использование ингибиторов мнк для борьбы с болезнью Альцгеймера
Последнее обновление: 2011-07-27
Частота использования: 1
Качество:
Английский
кристаллографическая структура белков mnk -1 и mnk -2
Французский
структура cristallographique des protéines mnk -1 и mnk -2
Последнее обновление: 2011-07-27
Частота использования: 1
Качество:
Английский
ИСПОЛЬЗОВАНИЕ ИНГИБИТОРОВ МНК ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ АЛЬЦГЕЙМЕРА
Французский
UTILIZATION D’INHIBITEURS DE MNK POUR LE TRAITEMENT DE LA MALADIE D’ALZHEIMER
Последнее обновление: 2011-07-27
Частота использования: 1
Качество:
Получите лучший перевод с
4 401 923 520
человеческий вклад
Сейчас обращаются за помощью пользователи:
Мы используем файлы cookie, чтобы вам было удобнее.Продолжая посещать этот сайт, вы соглашаетесь на использование файлов cookie. Учить больше.
ОК
MNK — Стоимость акций Mallinckrodt Plc
На странице «Обзор цен» можно просмотреть снимок определенного символа. Цены в режиме реального времени предоставляются Cboe BZX Exchange на отдельных страницах котировок акций США. В часы работы рынка в режиме реального времени отображается цена Cboe BZX, а на странице обновляются новые торговые данные (на что указывает «мигание»). Объем всегда отражает консолидированные рынки.Если по символу есть сделки до или после рынка, эта информация также будет отражена вместе с последней ценой (ценой закрытия), полученной при обмене символа. Цены в реальном времени доступны в часы работы рынка (с 9:30 до 16:00 EST).
Примечание : В настоящее время на биржу Cboe BZX приходится примерно 11–12% всей торговли акциями США каждый день. В результате цены, отображаемые в реальном времени, могут иметь незначительные расхождения при сравнении информации с другими сайтами, предлагающими данные в реальном времени, или с брокерскими фирмами.Если вам требуются комплексные ставки / запросы / предложения в режиме реального времени, мы предлагаем безрисковую пробную версию одного из наших продуктов в режиме реального времени.
Сводная таблица котировок
Сводная таблица котировок отображает данные моментального снимка котировок. Когда доступно, информация о Bid и Ask от Cboe BZX Exchange обновляется по мере поступления новых данных. Объем также обновляется, но представляет собой отложенный консолидированный объем от обмена символа. Поля данных доски объявлений включают:
- Дневной максимум / минимум : самая высокая и самая низкая цена сделки для текущей торговой сессии.
- Открыть : Цена открытия для текущей торговой сессии отображается на дневной гистограмме максимума / минимума.
- Предыдущее закрытие : Цена закрытия предыдущей торговой сессии.
- Ставка: Последняя цена и размер ставки.
- Ask: Последняя цена аск и размер аска.
- Объем: Общее количество акций или контрактов, торгуемых за текущую торговую сессию.
- Средний объем : Среднее количество акций, проданных за последние 20 дней.
- Взвешенный альфа : показатель того, насколько акции или товары выросли или упали за год. Barchart берет этот альфа и взвешивает его, присваивая больший вес недавним действиям и меньший (коэффициент 0,5) активности в начале периода. Таким образом, взвешенная альфа — это показатель годового роста с упором на самую последнюю ценовую активность.
Снимок графика
Предоставляется эскиз дневного графика со ссылкой для открытия и настройки полноразмерного графика.
Barchart Technical Opinion
Виджет Barchart Technical Opinion показывает сегодняшнее общее мнение Barchart с общей информацией о том, как интерпретировать краткосрочные и долгосрочные сигналы. Уникальная функция Barchart.com — система Opinions анализирует акции или товары с помощью 13 популярных аналитических инструментов в краткосрочной, среднесрочной и долгосрочной перспективе. Результаты интерпретируются как сигналы покупки, продажи или удержания, каждый из которых имеет числовой рейтинг и суммируется с общим процентным рейтингом покупки или продажи. После каждого расчета программа присваивает исследованию стоимость покупки, продажи или удержания в зависимости от того, где находится цена по отношению к общей интерпретации исследования.Например, цена выше скользящей средней обычно считается восходящим трендом или покупкой.
Символу будет присвоен один из следующих общих рейтингов:
- Сильная покупка (больше, чем «покупка на 66%»)
- Купить (больше или равна «покупка на 33%» и меньше или равно «Покупать на 66%»)
- Слабая покупка («Покупать 0% через« Покупать 33% »)
- Держать
- Сильная продажа (больше, чем« Продажа на 66% »)
- Продажа (больше или равно «33% продажи» и меньше или равно «66% продажи»)
- Слабая продажа («0% продажи» через «33% продажи»)
Текущее значение индикатора 14-Day Stochastic также учитывается при интерпретации.Следующая информация появится при выполнении следующих условий:
- Если 14-дневный стохастик% K больше 90 и Общее мнение — Покупка, отобразится следующее: «Рынок находится в зоне сильной перекупленности. Остерегайтесь разворот тренда «.
- Если 14-дневный стохастик% K больше 80 и Общее мнение — Покупка, появится следующее сообщение: «Рынок приближается к зоне перекупленности. Будьте осторожны при развороте тренда».
- Если 14-дневный стохастик% K меньше 10 и общее мнение является продажей, отображается следующее: «Рынок находится в зоне сильной перепроданности.Остерегайтесь разворота тренда ».
- Если 14-дневный стохастик% K меньше 20 и общее мнение является продажей, отображается следующее:« Рынок приближается к зоне перепроданности. Следите за разворотом тренда ».
Сводка по бизнесу
Предоставляет общее описание бизнеса, проводимого этой компанией.
Ценовые показатели
В этом разделе показаны максимумы и минимумы за последние 1, 3 и 12 месяцев Щелкните ссылку «Подробнее», чтобы просмотреть полную страницу отчета об эффективности с развернутой исторической информацией.
Основы
- Для акций США и Канады на странице «Обзор» представлены основные статистические данные об основных показателях акций со ссылкой для получения дополнительных сведений.
- Рыночная капитализация : капитализация или рыночная стоимость акции — это просто рыночная стоимость всех выпущенных акций. Он рассчитывается путем умножения рыночной цены на количество акций в обращении. Например, публичная компания с 10 миллионами акций, которые торгуются по 10 долларов каждая, будет иметь рыночную капитализацию в 100 миллионов долларов.
- Акции в обращении : Обыкновенные акции, находящиеся в обращении, по данным компании 10-Q или 10-K.
- Годовой объем продаж : Годовой объем продаж, выраженный в миллионах долларов.
- Годовой доход : Годовой чистый доход, выраженный в миллионах долларов.
- 60-месячная бета : коэффициент, который измеряет волатильность доходности акции относительно рынка (S&P 500). Он основан на 60-месячной исторической регрессии доходности акций на доходность S&P 500.
- Цена / Продажи : Последняя цена закрытия, деленная на доход / объем продаж на акцию за последние 12 месяцев.
- Цена / денежный поток : последняя цена закрытия, деленная на доход / денежный поток за последние 12 месяцев на акцию.
- Цена за книгу : Финансовый коэффициент, используемый для сравнения текущей рыночной цены компании с ее балансовой стоимостью.
- Цена / прибыль : последняя цена закрытия, деленная на прибыль на акцию за последние 12 месяцев.Компании с отрицательной прибылью получают «NE».
- Прибыль на акцию : Промежуточная прибыль на акцию за 12 месяцев от всех операций представляет собой чистую прибыль после всех расходов, деленную на средневзвешенное количество обыкновенных акций в обращении. Например, если компания имеет 10 миллионов долларов чистой прибыли и 10 миллионов выпущенных акций, то ее прибыль на акцию составляет 1 доллар.
- Последняя прибыль : сумма последней прибыли на акцию, выплаченной акционерам, и дата выплаты.Самые последние показатели прибыли основаны на доходах не по GAAP от продолжающейся деятельности.
- Дата следующего отчета : Дата следующего отчетного отчета.
- Годовые дивиденды и доходность : Указанная годовая ставка дивидендов и доходность, рассчитанные на основе последних дивидендов. Дивидендная ставка — это часть прибыли компании, выплачиваемая акционерам, выраженная в долларах, которые получает каждая акция (дивиденды на акцию). Доходность — это сумма дивидендов, выплачиваемых на акцию, деленная на цену закрытия.
- Самый последний дивиденд: Самый последний выплаченный дивиденд и самая последняя дата выплаты дивидендов.
- Сектора : ссылки на отраслевые группы и / или коды SIC, в которых находится товар.
Обзор опционов
Выделяет важную сводную статистику опционов, которая дает перспективные индикаторы настроений инвесторов.
- Подразумеваемая волатильность : Средняя подразумеваемая волатильность (IV) ближайшего месячного опционного контракта.IV — это прогнозируемый прогноз вероятности изменения цены базового актива, где более высокий IV означает, что рынок ожидает значительного движения цены, а более низкий IV означает, что рынок ожидает, что цена базового актива останется в пределах текущего торгового диапазона.
- 20-дневная историческая волатильность : среднее отклонение от средней цены за последние 20 дней. Историческая волатильность — это показатель того, насколько быстро базовая ценная бумага менялась в цене во времени.
- Процентиль IV : процент дней с закрытием IV ниже текущего значения IV по сравнению с предыдущим годом. Высокий процентиль IV означает, что текущий IV находится на более высоком уровне, чем в течение большей части прошлого года. Это может произойти после периода значительного движения цены, и высокий процентиль IV часто может предсказывать грядущий разворот цены на рынке.
- IV Рейтинг : текущий IV по сравнению с максимальным и самым низким значениями за последний год. Если рейтинг IV равен 100%, это означает, что IV находится на самом высоком уровне за последний год и может означать, что рынок перекуплен.
- IV High : самое высокое значение IV за последний год и дату, когда это произошло.
- Низкое значение IV : самое низкое значение IV за последний год и дату, когда это произошло.
- Соотношение объема пут / колл : общее соотношение объема пут / колл для всех опционных контрактов (для всех дат истечения). Высокое соотношение пут / колл может означать, что рынок перепродан, поскольку все больше трейдеров покупают путы, а не коллы, а низкое соотношение пут / колл может означать, что рынок перекуплен, поскольку все больше трейдеров покупают коллы, а не путы.
- Сегодняшний объем : общий объем всех опционных контрактов (по всем датам истечения), проданных в течение текущей сессии.
- Средний объем (30 дней) : средний объем для всех опционных контрактов (по всем датам истечения) за последние 30 дней.
- Коэффициент OI пут / колл : Коэффициент открытого интереса пут / колл для всех опционных контрактов (для всех дат истечения).
- Сегодняшний открытый интерес : общая сумма открытого интереса по всем опционным контрактам (за все даты истечения срока).
- Open Int (30-дневный) : Средняя общая сумма открытого интереса по всем опционным контрактам (по всем датам истечения) за последние 30 дней.
Связанные акции
Для сравнения найдите информацию о других символах, содержащихся в том же секторе.
Самые свежие новости
Просмотрите последние главные новости от Associated Press или Canadian Press (в зависимости от вашего выбора рынка).
Ингибирование MNK делает KRAS-мутантный колоректальный рак чувствительным к mTORC1 Ингибирование путем снижения фосфорилирования eIF4E и экспрессии c-MYC
Введение
Геномное секвенирование всесторонне охарактеризовало мутационный ландшафт колоректального рака (1, 2), обнаружив активирующие мутации в Протоонкоген KRAS у> 40% пациентов.Онкогенные мутации в KRAS приводят к конститутивной активации пролиферативных сигнальных каскадов, стимулируя прогрессирование опухоли и придавая устойчивость к стандартным методам лечения первой линии. Терапия, направленная на KRAS или его нижестоящие эффекторные пути, не реализовалась в клинике (3), хотя соединение, специфически нацеленное на мутантный KRAS G12C , недавно продемонстрировало эффективность при множественных солидных опухолях (4). Учитывая связь аберраций пути KRAS с резистентностью к терапии и рецидивом заболевания при колоректальном раке, а также столь же мрачные перспективы для пациентов с мутантом KRAS с другими показаниями, срочно необходимы новые стратегии лечения.
Инактивация опухолевого супрессора APC, которая приводит к нарушению регуляции пропролиферативной передачи сигналов WNT, является ключевым инициирующим событием в формировании аденомы кишечника. Ранее мы показали, что энтероциты с дефицитом Apc зависят от mTORC1, что способствует удлинению трансляции, сенсибилизируя аденомы с дефицитом Apc к ингибитору mTORC1 рапамицину (5). Это было подтверждено клинически, когда рапамицин регрессировал полипами толстой кишки у пациентов с семейным аденоматозным полипозом (FAP), предрасположенным к образованию аденомы из-за мутации APC зародышевой линии (6).Несмотря на этот успех в аденомах с дефицитом APC , пациенты со спорадическим колоректальным раком не могут получить пользу от рапалогов (рапамицина и его аналогов), а пациенты с онкогенными мутациями KRAS работают еще хуже (7–9). Важно отметить, что мыши, несущие опухоли толстой кишки с дефицитом Apc , продемонстрировали значительную регрессию с рапамицином, тогда как аналоги с потерей Apc и активацией KRAS не ответили (10). Эти соображения подчеркивают необходимость выявления терапевтических уязвимостей мутантных опухолей KRAS , которые могут взаимодействовать с рапалогами.
Колоректальный рак подразделяется на четыре согласованных молекулярных подтипа (CMS; ссылка 1). CMS1 обладает высокой степенью иммунной инфильтрации, часто встречаются мутации восстановления несоответствия. CMS4 сильно стромален и имеет наихудший прогноз. CMS2 и 3 характеризуются низкой инфильтрацией и в основном состоят из эпителиальных опухолевых клеток. CMS2 имеет высокие сигнатуры WNT и MYC, а CMS3 обогащен мутацией KRAS и обнаруживает метаболические изменения. Шкала микросреды Глазго (GMS) разделяет колоректальный рак по иммунной и стромальной инфильтрации (11), при этом GMS0 инфильтрируется иммунитетом (примерно коррелирует с CMS1), GMS2 является высоко стромальным с худшим прогнозом (сродни CMS4), а GMS1 — наименьшим. проникли (в соответствии с CMS2 / 3).Иммунологически редкие опухоли в CMS2 / 3 и GMS1 не реагируют на ингибирование контрольных точек и, следовательно, требуют новых терапевтических средств, направленных на ключевые внутренние пути раковых клеток.
При раке нарушение регуляции трансляции мРНК лежит в основе увеличенного синтеза белка в массе, который поддерживает повышенную скорость клеточной пролиферации, наряду с усилением экспрессии онкогена посредством преимущественного рекрутирования мРНК на рибосому (12). Фосфорилирование фактора инициации трансляции eIF4E по серину 209 (S209) находится ниже многих онкогенных путей; KRAS может передавать сигналы eIF4E через MAPK или p38 (13–16).Оба эти пути сходятся на MAP-киназа-взаимодействующих серин / треонин-протеинкиназ MNK1 и MNK2, которые, в свою очередь, являются единственными киназами, которые фосфорилируют eIF4E по S209 (17, 18). MNK-опосредованное фосфорилирование eIF4E участвует в твердом и жидком раке (19-25).
Делеция Mnk s допускается с точки зрения развития и не вызывает побочных эффектов у взрослых мышей (17), как и экспрессия нефосфорилируемого точечного мутанта S209A eIF4E (19). Переносимость и эффективность генетической делеции MNK в многочисленных моделях побудили к разработке ингибиторов MNK.Первым среди них является eFT508, мощный ингибитор с превосходной биодоступностью при пероральном приеме и фармакокинетикой, который перешел во II фазу клинических испытаний (24, 26). Механически фосфо-eIF4E (P-eIF4E) управляет программой онкогенной трансляции через набор «P-eIF4E-зависимых» транскриптов, включая C-myc, Cyclin D1, Cyclin E и Mcl1 (22, 23) . Как P-eIF4E специфически способствует трансляции онкогенных мРНК, остается неясным. Фактически, фосфорилирование серина 209 не требуется для продолжения трансляции и снижает сродство eIF4E к мРНК (27, 28).На сегодняшний день повышенная трансляция этих пролиферативных транскриптов коррелирует только с P-eIF4E. Остается немного данных, прямо демонстрирующих важность этих «P-eIF4E-зависимых» мРНК для фенотипа опухоли. Этот недостаток понимания ограничивает терапевтическое использование этого пути.
Здесь мы анализируем роль онкогенной мутации Kras на синтез белка и онкогенез в кишечнике. Используя генно-инженерные модели мышей (GEMM), мы демонстрируем, что активация KRAS усиливает синтез белка и стимулирует пролиферацию.Мы идентифицируем повышающую регуляцию инициации трансляции ниже по течению от KRAS, что совпадает с усилением передачи сигналов MNK / eIF4E. KRAS отменяет передачу сигналов от mTORC1, делая пролиферацию устойчивой к рапамицину и изменяя молекулярный режим рапамицина in vivo . Одного генетического или фармакологического нарушения оси MNK / eIF4E недостаточно для подавления онкогенеза. Однако совместное нацеливание пути с ингибированием mTORC1 с использованием рапамицина показывает значительное преимущество на множественных моделях органоидов на мышах и людях.Мы наблюдаем, что это включает подавление общего синтеза белка рапамицином и регуляцию экспрессии c-MYC с помощью P-eIF4E. Мы также показываем, что биомаркеры этих путей одновременно встречаются у пациентов с колоректальным раком и коррелируют с плохой выживаемостью у пациентов с опухолями GMS1, что составляет в общей сложности каждого пятого пациента. В совокупности наша работа определяет роль онкогенных KRAS в управлении синтезом белка и раскрывает уязвимость, поддающуюся лекарственному воздействию, ниже по течению от KRAS с терапевтическим потенциалом для лечения множества раковых заболеваний.
Результаты
Мутант
Kras придает устойчивость к рапамицину
Учитывая, что мутации KRAS коррелируют с устойчивостью к рапамицину, мы сначала исследовали влияние рапамицина после онкогенной мутации Kras . Это было достигнуто за счет создания мышей VillinCre ER Apc fl / fl Kras G12D / + (обозначаемых как краткосрочные APC KRAS), чьи ответы сравнивали с VillinCre ER Apc fl. / fl мышей (называемых APC short-term), экспрессирующих Kras дикого типа.После индукции внутрибрюшинной инъекцией тамоксифена у этих мышей наблюдается гиперпролиферация кишечника. Гиперпролиферация в кишечнике VillinCre ER Apc fl / fl снижается при лечении рапамицином из-за подавления гиперактивной передачи сигналов mTORC1 (рис. 1A), что согласуется с нашей предыдущей публикацией (5). Для сравнения, обработка рапамицином мышей VillinCre ER Apc fl / fl Kras G12D / + оказала лишь умеренное влияние на пролиферацию (рис.1А).
Рисунок 1.
Онкогенный KRAS приводит к агрессивным моделям опухолей, устойчивых к рапамицину, с дефицитом Apc . A, Слева: включение BrdU в тонкую кишку обработанных носителем или рапамицином Apc fl / fl и Apc fl / fl Kras G12D / + краткосрочная модель мышей в возрасте 4 лет и старше. Через 3 дня после индукции соответственно. Мышам вводили BrdU за 2 часа до отбора проб. Подсчитывали ≥25 полукрипт на мышь, каждая точка представляла биологическую копию. P значения взяты из односторонних тестов ANOVA множественных сравнений Тьюки. Справа на микрофотографиях показано окрашивание гематоксилином и эозином (H&E) и BrdU. Красной линией отмечена протяженность пролиферативной зоны. B, Вверху, схематическое изображение графика лечения. Внизу: выживаемость Apc fl / + Kras G12D / + мышей модели опухоли, получавших рапамицин или носитель ежедневно через 5 дней после индукции. C, Вверху, схематическое изображение графика лечения. Включение BrdU оценивали в опухолях тонкой кишки у мышей Apc 1322T / + , получавших рапамицин в течение 10 дней, с активацией Rosa CreER и без нее через 7 дней после начала активации рапамицина.Средний процент BrdU-положительности оценивали в ≥2 аденомах на мышь, где каждая точка представляла биологический повтор. P значения взяты из односторонних тестов ANOVA множественных сравнений Тьюки. Показаны репрезентативные изображения опухолей (пунктирные линии), окрашенные на антитело BrdU. Для сравнения показана пролиферация у необработанных мышей (левая полоса). Все данные представлены как среднее ± SEM. Масштабные линейки, 50 мкм. См. Также дополнительный рис. S1.
Параллельно мы создали мышей VillinCre ER Apc fl / + Kras G12D / + (т.е.е., теперь гетерозиготные по аллелю Apc flox — называемые мышами модели опухоли APC KRAS), у которых развиваются аденомы кишечника при потере второй копии Apc . Их выдерживали до появления признаков кишечного заболевания и сравнивали с когортой мышей VillinCre ER Apc fl / + (мыши с моделью опухоли APC). Экспрессия онкогенного KRAS снизила латентность нашей модели опухоли с 227 до 63 дней (дополнительный рис. S1A), что согласуется с предыдущими сообщениями (29, 30).У мышей, экспрессирующих онкогенный KRAS, наблюдается больше аденом толстой кишки, с меньшим увеличением аденом тонкой кишки (дополнительный рисунок S1B). Используя эту модель, мы проверили влияние рапамицина на инициирование и прогрессирование опухоли путем обработки мышей модели опухоли APC KRAS рапамицином или носителем через 5 дней после индукции, не увидев разницы в выживаемости (фиг. 1B). Ранее мы продемонстрировали полное подавление модели старения APC обработкой рапамицином (5). В совокупности эти данные указывают на то, что рапамицин не ингибирует образование или рост аденом в присутствии онкогенной мутации Kras G12D .Недавняя литература предполагает, что специфические мутации KRAS обладают разной способностью к трансформации, в зависимости от задействования нижестоящих эффекторных путей. При колоректальном раке KRAS A146T может вызывать устойчивость к EGFR, но коррелирует с лучшей выживаемостью, чем KRAS G12D , и менее эффективен при взаимодействии с потерей Apc на моделях мышей (31). В отличие от устойчивости к EGFR, мы обнаружили, что экспрессия KRAS A146T недостаточна для придания внутренней устойчивости рапамицину in vivo (дополнительный рис.S1C).
Затем мы использовали другую модель опухоли, мутантную мышь Apc 1322T / + , у которой развиваются аденомы кишечника, аналогичные пациентам с FAP. Лечение симптоматических мышей Apc 1322T / + , дикого типа для Kras , рапамицином в течение 10 дней приводило к снижению пролиферации сформировавшихся аденом (рис. 1С). Конструирование этих мышей для экспрессии онкогенного KRAS G12D в течение последних трех дней лечения рапамицином полностью изменило подавление пролиферации (рис.1С; Дополнительный рис. S1D), показывающий, что острая активация KRAS G12D достаточна для обеспечения быстрой и полной устойчивости к рапамицину.
Мутант
Kras способствует инициации трансляции и изменяет молекулярные эффекты рапамицина
Затем мы проанализировали ex vivo органоидов, образовавшихся в тонком кишечнике мышей APC или APC KRAS с коротким сроком действия. Эти органоиды воспроизводили фенотип in vivo , при этом органоиды APC давали сниженные метаболические показатели, используемые в качестве суррогата пролиферации после обработки рапамицином, тогда как органоиды APC KRAS не были затронуты (рис.2А; Дополнительный рис. S2A). Органоиды АРС, обработанные рапамицином, демонстрируют заметное снижение синтеза белка, измеренное по включению метионина 35 S (фиг. 2B). Органоиды APC KRAS также демонстрируют снижение синтеза белка на 25% (рис. 2A и B).
Рисунок 2.
Трансляция и пролиферация не связаны после мутации Kras . A, Рост органоидов Apc fl / fl или Apc fl / fl Kras G12D / + после 30-часовой обработки рапамицином 1 мкмоль / л по сравнению с органоидами, обработанными носителем.Были выполнены три технических повтора и усреднены, чтобы показать биологические повторы, как показано на графике. Значения P взяты из парных тестов Student t . Относительный рост для необработанных органоидов Apc fl / fl и Apc fl / fl Kras G12D / + установлен равным 0. B, Синтез белка в органоидах тонкого кишечника указанных генотипов после 6 часов лечения рапамицином 250 нмоль / л или носителем. Значения взяты из трех независимых биологических повторностей ± SEM, представленные относительно Apc fl / fl обработанных носителем органоидов.Значения P взяты из парных тестов Стьюдента t или теста Манна – Уитни. См. Также дополнительный рис. S2.
Чтобы получить представление о механизме действия рапамицина, мы проанализировали скорость инициации и удлинения трансляции с помощью ультрацентрифугирования плотности сахарозы и анализов стока харрингтонина. Модель APC KRAS демонстрирует больше полисом, но не отличается по скорости элонгации трансляции по сравнению с моделью APC (дополнительный рис. S2B и S2C), что указывает на повышенную инициацию трансляции, управляемую KRAS G12D .После обработки рапамицином клетки, дефицитные по Apc , реагируют замедлением удлинения и сохранением большего количества рибосом в полисомах (дополнительные рисунки S2B и S2C). Напротив, в ответ на рапамицин клетки APC KRAS загружают меньше рибосом в свои полисомы без изменения скорости удлинения, что указывает на нарушение инициации. Это демонстрирует измененный способ действия рапамицина — от ингибитора элонгации в Apc -дефицитных клетках до ингибитора инициации после мутации Kras .
Удлинение трансляции замедляется после лечения рапамицином в модели APC из-за увеличения P-eEF2 (5). В соответствии с отсутствием эффекта на удлинение трансляции в кишечнике APC KRAS, обработка рапамицином не влияла на P-eEF2 в этом генотипе (дополнительные рисунки S2D и S2E). Рапамицин остается эффективным ингибитором передачи сигналов S6K ниже mTORC1, при этом фосфорилирование его субстрата P-RPS6 S240 / 4 слегка подавляется в модели APC KRAS, тогда как P-4E-BP1 T37 / 46 значительно снижается под действием рапамицина.Следовательно, активация KRAS заменяет регуляцию P-eEF2 с помощью mTORC1, так что ингибирование mTORC1 больше не влияет на уровни P-eEF2. mTORC1 / S6K может также сигнализировать факторам инициации eIF4B и eIF4G (32–34). Мы обнаружили, что эти факторы инициации чувствительны к рапамицину в органоидах APC и APC KRAS, демонстрируя функциональную связь между mTORC1 и инициацией, которая, а также снижение P-4E-BP1 может объяснить, как рапамицин подавляет трансляцию в модели APC KRAS (дополнительная информация Инжир.S2B, S2D и S2E).
Из этого мы заключаем, что активация KRAS с делецией Apc усиливает синтез белка, способствуя инициации трансляции. Мы также наблюдаем функциональное разделение синтеза белка с пролиферацией, при этом обработка рапамицином органоидов APC KRAS приводит к подавлению синтеза белка без влияния на пролиферацию. Таким образом, мы пришли к выводу, что для успешного нацеливания трансляции в этой модели нам необходимо либо подавить перевод дальше, либо выборочно подавить трансляцию пропролиферативных сообщений.Чтобы исследовать, мы сосредоточились на инициации трансляции, учитывая ее важность после активации KRAS.
P-eIF4E усилен
Kras G12D Мутация
Известно, что фосфорилирование eIF4E происходит ниже по ходу передачи сигналов KRAS (35). Мы наблюдали увеличение P-eIF4E в краткосрочном кишечнике APC KRAS и в аденомах толстой кишки из опухолевой модели APC KRAS по сравнению с аналогами APC (фиг. 3A; дополнительный рисунок S3A). Соответственно, вестерн-блоттинг показал ступенчатое увеличение P-eIF4E до культур APC, а затем и APC KRAS по сравнению с органоидами дикого типа (дополнительный рис.S3B). Интересно, что P-eIF4E был ниже в кишечнике, экспрессирующем Kras A146T / + (дополнительный рисунок S3C), что согласуется с ответом на лечение рапамицином, наблюдаемым в опухолях с этим аллелем Kras (дополнительный рисунок S1C). Онкогенный BRAF также способен передавать сигналы через те же пути к P-eIF4E. P-eIF4E был вариабельным в кишечнике мышей Braf V600E / + (дополнительный рис. S3C), что позволяет предположить, что этот путь может играть роль у ~ 10% пациентов с этими мутациями.
Рисунок 3.
P-eIF4E увеличивается за счет мутации Kras и действует синергетически с mTORC1, способствуя пролиферации. A, ИГХ окрашивание на P-eIF4E в краткосрочной модели (2 панели слева) и аденомы толстой кишки на модели опухоли (2 панели справа) из Apc fl / fl или + и Apc fl / fl или + Kras G12D / + мышей. Пунктирная линия обозначает границу аденомы. Изображения представляют ≥3 биологических повторов. B, Вверху, схематическое изображение графика лечения. Apc fl / + Kras G12D / + и Apc fl / + Kras G12D / + Eif4e S209A / S209A Мышам с опухолевой моделью было разрешено развить симптомы опухолей кишечника, леченных рапамицином в течение 5 дней, а затем через 2 часа после введения BrdU. Внизу: включение BrdU в клетки эпителиальной аденомы толстой кишки, количественно определенное для минимум четырех опухолей на мышь, причем каждая точка представляет собой биологическую копию. P значения взяты из односторонних тестов ANOVA множественных сравнений Тьюки. C , внизу, количественная оценка включения BrdU в Apc fl / fl Kras G12D / + Eif4e S209A / S209A краткосрочная модель мышей, получавших рапамицин или носитель. Вверху: репрезентативное окрашивание H&E, BrdU и P-eIF4E для каждого лечения, с красной полосой, показывающей степень распространения. Для каждой мыши было определено ≥25 полукрипт, среднее значение для каждой мыши нанесено на график. P Значение взято из теста Манна – Уитни. D, Типичные изображения окрашивания для P-eIF4E и P-RPS6, каждое из которых дуплексировано с цитокератином из ТМА пациентов с колоректальным раком.Для каждого показаны примеры сильного и слабого окрашивания. E, Корреляция между P-eIF4E и P-RPS6 из TMA, показанная в D . Значимость проверялась с помощью теста X 2 . Все данные представлены как среднее ± SEM. Масштабные линейки, 50 мкм. См. Также дополнительный рис. S3.
Затем мы попытались ослабить P-eIF4E и наблюдали его влияние на онкогенез в моделях, управляемых Kras G12D . Генетическое изменение серина 209 eIF4E на аланин предотвращает восходящую передачу сигналов от MNK и переносится in vivo (19).Таким образом, мы создали мышей VillinCre ER Apc fl / (fl или +) Kras G12D / + Eif4e S209A / S209A мышей, которые имеют гомозиготный нокаут eIF4E S209A KRAS eIFC краткосрочные или опухолевые модели). Начало кишечной неоплазии существенно не отличалось в опухолевой модели APC KRAS eIF4E KI по сравнению с контрольными APC KRAS, и у мышей развивалась аналогичная опухолевая нагрузка (дополнительный рисунок S3D).
Учитывая, что рапамицин подавляет синтез белка посредством инициации трансляции в модели APC KRAS, мы пришли к выводу, что сочетание рапамицина с нацеливанием на P-eIF4E может быть полезным.Обработка рапамицином мышей APC KRAS eIF4E KI с ранней временной точки (5 дней) привела к увеличению выживаемости на 65% по сравнению с обработанными рапамицином мышами APC KRAS (дополнительный рисунок S3E). Затем мышей APC KRAS eIF4E KI лечили рапамицином в более поздний момент времени, когда у них были аденомы кишечника. У этих животных наблюдалось значительное снижение пролиферации аденомы после 5 дней лечения (фиг. 3B) и 4-кратное увеличение выживаемости (дополнительные фиг. S3F и S3G). Мыши APC KRAS (дикий тип для Eif4e ), получавшие рапамицин на той же стадии кишечного заболевания, не получали пользы от рапамицина (рис.3Б). В соответствии с этим мы наблюдали снижение кишечной пролиферации на краткосрочной модели APC KRAS eIF4E KI после лечения рапамицином (рис. 3C) и в органоидах ex vivo от этих мышей при лечении рапамицином (дополнительный рис. S3H) . P-eIF4E отсутствовал у мышей, несущих мутацию S209A (рис. 3C). Рапамицин не влиял на P-eIF4E в краткосрочной модели APC KRAS Eif4e дикого типа (дополнительный рисунок S3A и S3I), тем самым исключая этот потенциальный механизм нарушения P-eIF4E, повышающего чувствительность к рапамицину.
P-RPS6 S240 / 4 и P-eIF4E S209 одновременно встречаются у пациентов с колоректальным раком
Эффективность нацеливания на P-eIF4E в сочетании с ингибированием mTORC1 указывает на важность этих путей при колоректальном раке. Чтобы выяснить клиническую важность этих путей, мы окрасили тканевый микрочип (ТМА), содержащий опухоли от 192 пациентов, на P-eIF4E S209 и P-RPS6 S240 / 4, последний как показания активности mTORC1 (рис. 3D). Почти 50% (80/170) пациентов имели высокие уровни как P-eIF4E, так и P-RPS6, что способствовало значительной положительной корреляции между ними (рис.3E). Таким образом, высокая активность MNK и mTORC1 наблюдается у значительной части пациентов с колоректальным раком, что позволяет идентифицировать популяцию пациентов, которым может быть полезен котаргетинг.
Mnk1 / 2 Делеция объединяется с рапамицином для подавления KRAS-управляемой пролиферации
Увеличение P-eIF4E указывает на усиление передачи сигналов через MNK. Таким образом, мы создали VillinCre ER Apc fl / (fl или +) Kras G12D / + Mnk1 — / — Mnk2 — / — мышей (называемых APC KRAS MNK1 / 2 KO short- term или опухолевые модели), которые являются нокаутом зародышевой линии (KO) для Mnk1 и Mnk2 (17), чтобы оценить важность этих киназ.Параллельно мы сгенерировали VillinCre ER Apc fl / (fl или +) Kras G12D / + с одним выбиванием Mnk1 или Mnk2 . Делеция Mnk2 оказала большее влияние на P-eIF4E, чем делеция Mnk1 (фиг. 4A), что согласуется с более высокой экспрессией Mnk2 в кишечнике мыши (дополнительная фиг. S4A). Mnk1 и Mnk2 экспрессируются по всему эпителию кишечника мышей APC и APC KRAS (рис.4Б). Экспрессия Mnk2 , по-видимому, повышена в APC KRAS по сравнению с APC в кишечнике, что подтверждается кПЦР для каждого транскрипта Mnk (фиг. 4C). BaseScope также подтвердил отсутствие транскриптов Mnk1 и Mnk2 у мышей APC KRAS с нокаутами Mnk (дополнительный рисунок S4B). Удаление как Mnk1 , так и Mnk2 уничтожило P-eIF4E in vivo (фиг. 4A), как это произошло с органоидами (дополнительный рисунок S4C). Хотя органоиды APC KRAS, несущие нокауты Mnk1 или Mnk2 , демонстрировали пониженные уровни P-eIF4E по сравнению с их аналогами дикого типа, делеция Mnk1 оказывала большее влияние на P-eIF4E в культурах органоидов, чем в экспериментах in vivo .Причины этого неясны, но могут быть связаны с большей ролью киназы в культуре или отсутствием компенсации со стороны MNK2.
Рисунок 4. Делеция
Mnk подавляет пролиферацию опухоли, вызванную мутантом KRAS , в сочетании с лечением рапамицином. A, IHC для P-eIF4E в тонком кишечнике Apc fl / fl Kras G12D / + краткосрочная модель мышей с делецией Mnk1, Mnk2 или обоих Mnk1 и Mnk2 .Справа, количественная оценка этого окрашивания P-eIF4E. B, BaseScope анализ экспрессии Mnk1, и Mnk2 в Apc fl / fl и Apc fl / fl Kras G12D / + тонкой кишки. C, КПЦР для Mnk1 и Mnk2 в Apc fl / fl и Apc fl / fl Kras G12D / + тонкой кишки. Уровни выражены относительно экспрессии мРНК Actb . D, Вверху, схематическое изображение графика лечения. Apc fl / + Kras G12D / + и Apc fl / + Kras G12D / + Mnk1 — / — Mnk2 — / — Мыши с опухолью стареют до появления симптомов кишечной опухолей, а затем лечили рапамицином или носителем до конечной точки. Выживаемость отображается как количество дней лечения рапамицином или носителем. P Значение получено из логарифмического теста Мантела – Кокса. E, включение BrdU в тонком кишечнике Apc fl / fl Kras G12D / + и Apc fl / fl Kras G12D / + Mnk1 — / — Mnk2 — / — краткосрочная модель мышей, получавших рапамицин и без него. Apc fl / fl Kras G12D / + Данные воспроизведены с рис. 1A для простоты сравнения. Для каждой мыши было определено ≥25 полукрипт, среднее значение для каждой мыши показано на представленном рисунке. P значения взяты из односторонних тестов ANOVA множественных сравнений Тьюки. Горизонтальная линия представляет среднее значение для мышей Apc fl / fl Kras G12D / + . F, График выживаемости после ортотопической индукции в дистальном отделе толстой кишки Apc fl / fl Kras G12D / + ( n = 3) или Apc fl / fl Kras G12D / + Mnk1 — / — Mnk2 — / — мышей ( n = 4).Когорта мышей Apc fl / fl Kras G12D / + Mnk1 — / — Mnk2 — / — мышей также лечили рапамицином с 16-го дня ( n = 4), обозначено вертикальной штриховой линией. линия. Полипы толстой кишки были визуализированы с помощью колоноскопии в указанные дни. P Значение получено из логарифмического теста Мантела – Кокса. Мышь Apc fl / fl Kras G12D / + Mnk1 — / — Mnk2 — / — на рапамицине была подвергнута цензуре через 100 дней после индукции, не связанной с процедурой, и две были подвергнуты цензуре в конце исследования. через 202 дня после индукции.Все данные представлены как среднее ± SEM. Масштабные линейки, 50 мкм. См. Также дополнительный рис. S4.
Одинарная или двойная делеция Mnk s в модели опухоли APC KRAS не влияла на выживаемость по сравнению с мышами, экспрессирующими оба Mnk (дополнительный рисунок S4D). Мыши APC KRAS MNK1 / 2 KO, которым вводили рапамицин при проявлении симптомов поздней стадии кишечного заболевания, выживали в 10 раз дольше, чем мыши APC KRAS MNK1 / 2 KO, обработанные носителем, или мыши APC KRAS, обработанные рапамицином (рис.4D). Сходным образом, в краткосрочной модели, APC KRAS MNK1 / 2 KOs показали частичное подавление пролиферации по сравнению с контрольными APC KRAS, а в сочетании с рапамицином продемонстрировали сниженную гиперпролиферацию (фиг. 4E). Мы наблюдали ступенчатое снижение эффекта комбинации с рапамицином в моделях APC KRAS с нокаутом Mnk1 с последующим нокаутом Mnk2 и, наконец, с нокаутом Mnk1 и Mnk2 как в опухолевой, так и в краткосрочной моделях (дополнительный рис. .S4E – S4H). Эта дозозависимая природа нокаута Mnk обнадеживает для нацеливания на лекарства, предлагая широкое терапевтическое окно.
Затем мы использовали ортотопический подход, согласно которому VillinCre ER Apc fl / fl Kras G12D / + или VillinCre ER Apc fl / fl Kras G12D / + Mnk / — Mnk2 — / — Мыши получили местную инъекцию тамоксифена в подслизистую основу дистального отдела толстой кишки, в результате чего образовался единственный полип толстой кишки, который можно продольно контролировать с помощью колоноскопии (36, 37).Мыши APC KRAS MNK1 / 2 KO, получавшие рапамицин, выживали значительно дольше, чем необработанные мыши APC KRAS или APC KRAS MNK1 / 2 KO (фиг. 4F; дополнительный фиг. S4I). Две из четырех мышей APC KRAS MNK1 / 2 KO выжили до 200 дней после индукции по сравнению со средней выживаемостью всего 29 дней в контроле APC KRAS. Таким образом, генетическое воздействие на путь MNK / eIF4E в сочетании с лечением рапамицином подавляет онкогенез. Затем мы исследовали механизм этого успешного лечения.
P-eIF4E и mTORC1 взаимодействуют для регуляции экспрессии c-MYC
Градиенты плотности сахарозы экстрактов из APC KRAS eIF4E KI и APC KRAS MNK1 / 2 KO мышей, модельных краткосрочных моделей, показывают, что рапамицин незначительно снижает количество полисомов в обоих случаях (дополнительная информация Инжир.S5A и S5B), как и у мышей APC KRAS, что указывает на то, что рапамицин остается ингибитором инициации трансляции. Скорость оттока рибосом не изменилась при обработке рапамицином органоидов APC KRAS eIF4E KI или APC KRAS MNK1 / 2 KO (дополнительные рисунки S5C и S5D). Следовательно, сенсибилизация к рапамицину у этих двух генотипов не связана с изменением способа его действия на ингибитор удлинения трансляции, наблюдаемого в моделях APC. Рапамицин является эффективным ингибитором синтеза белка для всех проанализированных генотипов (дополнительный рис.S5E), а генетическое нацеливание на Mnk или Eif4E не изменило действие рапамицина на фосфорилирование eEF2 или RPS6 S240 / 4 (дополнительный рисунок S5F).
Скорость синтеза белка была сопоставима между обработанными рапамицином органоидами APC KRAS, которые не подавлены, и обработанными рапамицином органоидами APC KRAS eIF4E KI или APC KRAS MNK1 / 2 KO, у которых метаболическая способность клеток была ослаблена (дополнительный рисунок S5E). Интересно, что эти данные также указывают на то, что в отсутствие рапамицина делеция Mnk снижает синтез белка.Подавление MNK редко оказывает большое влияние на глобальный синтез белка, хотя данные, аналогичные представленным здесь, были показаны в двух отдельных отчетах с использованием клеток рака простаты (38, 39). Следовательно, подавление массового синтеза белка не является механизмом эффективности котаргетинга MNK / eIF4E и mTORC1, а скорее всего зависит от снижения трансляции специфических транскриптов. Поэтому мы стремились идентифицировать мРНК, связанные общим eIF4E и P-eIF4E путем иммунопреципитации РНК (RIP) из органоидов APC KRAS.Образцы иммуноблоттинга из этих RIP показывают обогащение P-eIF4E и восстановление общего eIF4E (как фосфорилированных, так и нефосфорилированных форм) из параллельных eIF4E RIP (фиг. 5A). qPCR РНК, очищенной из этих RIPs, выявляет однородное связывание транскриптов общим eIF4E между предсказанными «P-eIF4E-зависимыми» и контрольными мРНК (фиг. 5B; ссылки 20, 22, 23). qPCR из P-eIF4E RIP показывает обогащение для «P-eIF4E-зависимых» сообщений по сравнению с контрольными транскриптами ActB и Gapdh , при этом транскрипт C-myc значительно обогащает P-eIF4E RIP по сравнению с Расшифровка Gapdh (рис.5Б). Точно так же RIP, выполненные с использованием экзогенного FLAG-eIF4E, экспрессированного в клетках HCT116, обнаружили значительное обогащение c-MYC среди других P-eIF4E-зависимых транскриптов, чем контрольный транскрипт TUBA1B (дополнительный рисунок S5G). Важно отметить, что экспрессия FLAG-eIF4E с мутацией S209A снижает обогащение c-MYC по сравнению с контрольными транскриптами (дополнительный рисунок S5G). Это указывает на то, что FLAG-eIF4E, который может быть фосфорилирован, легче ассоциирует с c-MYC , чем нефосфорилируемый FLAG-eIF4E.
Рисунок 5.
Трансляция C-myc требует P-eIF4E и активной передачи сигналов mTORC1. A, Проверка иммунопреципитации (IP) общего eIF4E или P-eIF4E с помощью вестерн-блоттинга (WB) для каждой соответствующей мишени. B, вверху, схематически показано, что суммарное антитело eIF4E осаждает как фосфорилированный, так и нефосфорилированный eIF4E, тогда как антитело P-S209 eIF4E специфически осаждает фосфорилированную форму. Внизу: количественная ПЦР для мРНК-мишеней с использованием РНК, элюированной из IP eIF4E, P-S209 eIF4E или контрольного преципитации IgG, которая не элюировала eIF4E.Данные представлены в виде линеаризованных значений Ct для IP РНК, вычтенных из значения Ct для входных данных. Серая линия на каждом графике — это значение для расшифровки Gapdh . Все данные являются средними для четырех биологических повторов. Значимость проверялась парным тестом Student t . C , IHC для c-MYC в аденомах из Apc fl / + Kras G12D / + or Apc fl / + Kras G12D / + Mnk1 — / — Mnk2 — / — мышей, получавших рапамицин или без него в течение 5 дней до отбора проб.Интенсивность окрашивания количественно оценивали с использованием H-показателей ядерной c-MYC, показанных на соседнем графике. Средний H-балл, по крайней мере, для семи аденом на мышь, каждая точка показывает биологическую копию. P значения взяты из односторонних тестов ANOVA множественных сравнений Тьюки. Все данные представлены как среднее ± SEM. Масштабные линейки, 50 мкм. См. Также дополнительный рис. S5.
C-myc особенно эффективен при колоректальном раке, что продемонстрировано эффектами его делеции в GEMM, дефицитных по Apc (40).Мы проанализировали экспрессию ядерного c-MYC в аденомах толстой кишки у мышей модели опухоли APC KRAS MNK1 / 2 KO, получавших рапамицин или без него, по сравнению с мышами APC KRAS. Ядерная экспрессия c-MYC была снижена в аденомах из APC KRAS MNK1 / 2 KOs, обработанных рапамицином (фиг. 5C), но относительно не изменилась в соответствующей краткосрочной модели (дополнительная фиг. S5H). Цитоплазматическая экспрессия c-MYC следовала аналогичному паттерну, но менее чем на половине уровней ядерного c-MYC (дополнительный рис. S5I). Вестерн-блоттинг лизатов органоидов APC KRAS MNK1 / 2 KOs также показал последовательное снижение экспрессии c-MYC после обработки рапамицином (дополнительный рис.S5J). Важно отметить, что экспрессия мРНК C-myc не изменялась присутствием или отсутствием МНК или обработкой рапамицином (дополнительный рисунок S5K). Следовательно, P-eIF4E обогащен мРНК C-myc , и, когда этот путь подавляется в сочетании с лечением рапамицином, экспрессия c-MYC падает.
Котаргетинг MNK и mTORC1 подавляет опухолевый генез, управляемый KRAS
Затем мы попытались обобщить наши генетические данные с помощью низкомолекулярных ингибиторов.Для ингибирования МНК мы использовали мощный ингибитор МНК eFT508 (24, 26). Обработка eFT508 краткосрочной модели APC KRAS существенно снижает P-eIF4E, но оказывает незначительное влияние на пролиферацию (фиг. 6A; дополнительная фиг. S6A). Комбинация eFT508 с рапамицином значительно снижала пролиферацию по сравнению с животными, получавшими оба носителя (фиг. 6A), до уровня, аналогичного KRAS MNK1 / 2 KO APC, обработанных рапамицином (фиг. 4E). eFT508 присутствовал в плазме более 3 нмоль / л и 300 нмоль / л в кишечнике через 2 часа после перорального желудочного зондирования, концентрации, которые не были изменены при одновременном дозировании с рапамицином (дополнительный рис.S6B). Аналогичный эффект был замечен в отношении МНК с помощью мультикиназного ингибитора мерестиниба (дополнительный рисунок S6C). Эффективность комбинации лекарственных средств не зависела от передачи сигнала к eEF2 через eEF2K, как показано подавлением пролиферации в комбинациях eFT508 / рапамицин или мерестиниб / рапамицин в VillinCre ER Apc fl / fl Kras G12D / + eEF2K D273A3 кишечник (дополнительные рисунки S6C и S6D; ссылка 41). При анализе в модели APC, зависящей от элонгации, этот аллель, инактивирующий eEF2K, полностью устраняет чувствительность к рапамицину (дополнительный рис.S6E). Это подтверждает, что рапамицин (и ингибирование MNK) не снижает пролиферацию через eEF2K-зависимое замедление элонгации. Комбинация рапамицина и eFT508 в этой модели APC (с диким типом Kras ) не оказывала дополнительного воздействия на пролиферацию, чем лечение одним рапамицином (дополнительный рисунок S6E).
Рисунок 6.
Нацеливание на MNK с помощью eFT508 в сочетании с рапамицином для подавления фенотипа APC KRAS. A, включение BrdU в Apc fl / fl Kras G12D / + краткосрочная модель мышей, получавших eFT508 или eFT508 в комбинации с рапамицином.Для каждой мыши было определено ≥25 полукрипт, среднее значение для каждой мыши показано на представленном рисунке. P Значение получено из односторонних тестов ANOVA множественных сравнений Тьюки. B, Apc fl / + Kras G12D / + мышей с моделью опухоли лечили только eFT508 или в комбинации с рапамицином в течение 5 дней после появления симптомов аденомы кишечника. BrdU вводили за 2 часа до отбора образцов, а затем количественно определяли процент BrdU-положительных клеток аденомы.На каждую мышь было проанализировано ≥5 аденом, а затем их среднее значение было нанесено на график. Каждая точка представляет собой биологическую копию. P Значение получено из односторонних тестов ANOVA множественных сравнений Тьюки. C, Микрофотографии, показывающие характерное окрашивание для включения BrdU и уровней P-eIF4E и c-MYC из B . D, Количественная оценка окрашивания c-MYC в C по H-баллу. P Значение получено из односторонних тестов ANOVA множественных сравнений Тьюки. E, Слева, схематическое изображение графика лечения. Справа: Apc fl / + Kras G12D / + мышей с моделью опухоли, обработанных как в B , и позволили стареть в процессе лечения до клинической конечной точки. P Значение получено из логарифмического теста Мантела – Кокса. F, Apc fl / fl Kras G12D / + органоидов обрабатывали eFT508 (30 нмоль / л) или eFT508 в сочетании с рапамицином (250 нмоль / л) в течение 24 часов, и изменение роста по сравнению с клетками, обработанными носителем.Каждая точка данных представляет собой независимую биологическую копию, которая представляет собой среднее трех технических повторений. Значение P взято из парного теста Student t . Все данные представлены как среднее ± SEM. Масштабные линейки, 50 мкм. См. Также дополнительный рис. S6.
eFT508 / рапамицин был эффективен в модели опухоли APC KRAS, значительно подавляя пролиферацию через 5 дней лечения с уменьшением P-eIF4E (фиг. 6B и C). Подавление пролиферации с помощью eFT508 / рапамицина в аденомах APC KRAS коррелировало со снижением ядерной и цитоплазматической экспрессии c-MYC (рис.6C и D; Дополнительный рис. S6F). Органоиды APC KRAS, обработанные комбинацией в течение 24 часов, также имели пониженную экспрессию c-MYC (дополнительный рисунок S6G). Лечение мышей с аденомами также значительно увеличивало выживаемость: животные жили до 120 дней после лечения по сравнению с менее чем 10 днями при лечении носителем (фиг. 6E). Комбинированное лечение было чрезвычайно хорошо переносимым, с отличной модуляцией биомаркеров, но без побочных эффектов на гомеостаз у мышей дикого типа (дополнительный рис.S6H). Экспрессия c-MYC также не изменилась при комбинированном лечении у мышей дикого типа, что указывает на то, что эта модуляция специфична для модели опухоли APC KRAS, а не более широкое влияние на экспрессию c-MYC. Обработка модели ортотопического полипа толстой кишки APC KRAS с помощью eFT508 / рапамицина привела к аналогичному увеличению выживаемости по сравнению с обработкой одним лекарственным средством (дополнительный рисунок S6I), а обработка органоидов APC KRAS рапамицином и eFT508 в комбинации снизила метаболическую активность клеток. , заменитель числа клеток, на ∼40% (рис.6F; Дополнительный S6J).
Механически обработка eFT508 была аналогична обработке с нокаутом Mnk . eFT508 подавлял синтез белка без дополнительного ингибирования при сочетании с рапамицином (дополнительный рисунок S6K). Следует отметить, что RIP, выполненные после обработки eFT508 органоидов APC KRAS, не показали изменения количества c-Myc , связанного общим eIF4E (дополнительный рисунок S6L). Таким образом, P-eIF4E не увеличивает привлечение eIF4E к мРНК, что согласуется с предыдущими сообщениями (27, 42), но, вероятно, способствует трансляции транскриптов при связывании.
Эффективность MNKi и рапамицина полностью зависит от регуляции c-MYC
Нацеливание на P-eIF4E в сочетании с лечением рапамицином последовательно приводит к снижению экспрессии ядерного белка c-MYC (фиг. 5C и 6D). Однако неясно, является ли это причиной или следствием снижения распространения. Чтобы проверить это, мы использовали индуцибельную кДНК C-MYC человека, лишенную 5′- или 3′-UTR, которой предшествовала Cre-активированная кассета LoxP-STOP-LoxP (lsl) в локусе Rosa26 ( R26-lsl -MYC ; исх.43). После рекомбинации две дополнительные копии c-MYC экспрессируются в кишечном эпителии одновременно с делецией Apc и активацией KRAS G12D . Эти мыши обозначаются как APC KRAS MYC.
Экспрессия дополнительного c-MYC имела незначительный эффект на пролиферацию в сформировавшихся аденомах (фиг. 7A) с аналогичными ядерными уровнями c-MYC между необработанными APC KRAS и APC KRAS MYC моделями мышей (фиг. 7B). Однако пролиферация аденом этих мышей APC KRAS MYC не пострадала при лечении eFT508 / рапамицином, в отличие от почти 50% снижения пролиферации в аденомах, обработанных APC KRAS (рис.7А). В соответствии с устойчивой пролиферацией, обработанные аденомы APC KRAS MYC показали более высокую экспрессию ядерного c-MYC по сравнению с обработанными аденомами APC KRAS (фиг. 7B). Органоиды APC KRAS MYC также показали повышенную среднюю экспрессию c-MYC после обработки eFT508 / рапамицином по сравнению с органоидами APC KRAS (дополнительный рисунок S6G). Цитоплазматический c-MYC следует той же схеме (дополнительный рисунок S7A). Кроме того, увеличение выживаемости, наблюдаемое с eFT508 / рапамицином у мышей модели опухоли APC KRAS (в среднем 49 дней), было полностью потеряно у мышей APC KRAS MYC (в среднем <4 дней; дополнительный рис.S7B), несмотря на то, что eFT508 подавляет P-eIF4E в аналогичной степени (дополнительный рисунок S7C). Трансген C-MYC дал аналогичные результаты на краткосрочной модели APC KRAS (рис. 7C) и культуре органоидов (рис. 7D), при этом пролиферация сравнима между мышами APC KRAS MYC или органоидами, обработанными eFT508 / рапамицином и носителем лечил БТР КРАС. В целом эти данные показывают, что подавление экспрессии белка c-MYC с помощью eFT508 / рапамицина полностью объясняет снижение пролиферации в модели APC KRAS.
Фигура 7.
Экспрессия человеческого трансгена C-MYC полностью отменяет положительный эффект лечения eFT508 / рапамицином in vivo . A, Top, индикатор выраженных вариантов c-MYC. Apc fl / + Kras G12D / + R26-lsl-MYC мышей с моделью опухоли лечили eFT508 в комбинации с рапамицином в течение 5 дней после появления симптомов аденомы кишечника. BrdU вводили за 2 часа до отбора образцов и количественно определяли процент BrdU-положительных клеток аденомы.Данные для контрольных мышей Apc fl / + Kras G12D / + воспроизведены с фиг. 6B для простоты сравнения. Процент положительности BrdU был усреднен по крайней мере в пяти аденомах на мышь, причем каждая точка представляла отдельную мышь. P значения взяты из односторонних тестов ANOVA множественных сравнений Тьюки. B, окрашивание c-MYC (внизу) и количественная оценка H-score (вверху) в аденомах из Apc fl / + Kras G12D / + и Apc fl / + Kras G12D / + Модель опухоли R26-lsl-MYC у мышей после 5 дней лечения eFT508 / рапамицином.H-показатель C-MYC был определен для ≥7 аденом на мышь, а затем среднее значение было нанесено на представленный график. P значения взяты из односторонних тестов ANOVA множественных сравнений Тьюки. C, Top, индикатор выраженных вариантов c-MYC. Включение BrdU в Apc fl / fl Kras G12D / + R26-lsl-MYC на краткосрочных моделях мышей, получавших eFT508 в комбинации с рапамицином. Значения включения BrdU для мышей Apc fl / fl Kras G12D / + , получавших eFT508 / рапамицин, воспроизведены на фиг.6А. ≥25 полукрипт оценивали на мышь, а затем наносили среднее значение в виде отдельной точки на графике. P значения взяты из односторонних тестов ANOVA множественных сравнений Тьюки. D, Top, индикатор выраженных вариантов c-MYC. Рост Apc fl / fl Kras G12D / + или Apc fl / fl Kras G12D / + R26-lsl-MYC органоидов, обработанных eFT508 (30 нмоль / л) и рапамицином (250 нмоль / л) в течение 24 часов по сравнению с ростом аналогов, обработанных носителем.Технические трижды усредняли для трех независимых линий органоидов, при этом эти средние значения представлены каждой точкой на графике. Значение P взято из парного теста Student t . E, график пути REACTOME, показывающий значительно положительно обогащенные пути в РНК-seq между обработанными рапамицином / eFT508 Apc fl / fl Kras G12D / + R26-lsl-MYC по сравнению с Apc fl / fl Kras G12D / + кишечник.Соотношение генов указывает на долю генов в пути, которые регулируются, с абсолютным числом, показанным размером круга. Цвет круга указывает на важность обогащения. F, Кривая выживаемости Каплана-Мейера для рака-специфической выживаемости для пациентов с GMS1, имеющих низкую окраску для одного или обоих из P-RPS6 S240 / 4 и P-eIF4E S209 ( n = 46) по сравнению с пациентами, имеющими высокую окраску для обоих маркеров ( n = 42). Средняя выживаемость аннотируется в каждой строке. Значимость определялась с помощью лог-рангового теста.Все данные представлены как среднее ± SEM. Масштабные линейки, 50 мкм. См. Также дополнительный рис. S7.
Обработка eFT508 / рапамицином подавляла синтез белка в клетках APC KRAS MYC в той же степени, что и органоиды APC KRAS (дополнительный рисунок S7D). Таким образом, повторная экспрессия c-MYC не вызывает пролиферацию путем простого восстановления синтеза белка. Мы предполагаем, что рапамицин подавляет глобальный синтез белка, создавая потребность в P-eIF4E для стимуляции экспрессии c-MYC. Чтобы проверить это, мы использовали три терапевтически релевантных агента, которые подавляют общий синтез белка, оксалиплатин, 5-фторурацил (5-FU) и рокагламид (44–46) вместо рапамицина, обнаружив, что органоиды APC KRAS были сенсибилизированы к лечению. с этими соединениями при совместном введении eFT508 (дополнительный рис.S7E). Эта сенсибилизация зависела от подавления c-MYC, поскольку органоиды APC KRAS MYC были невосприимчивы к комбинированной обработке eFT508. Таким образом, ингибирование MNK действует для подавления пролиферации в опухолевых клетках, где синтез белка ограничен, путем подавления экспрессии c-MYC.
c-MYC восстанавливает пролиферацию путем изменения транскрипции метаболических генов
Трансген c-MYC не может восстанавливать скорость синтеза белка после лечения eFT508 / рапамицином даже после 1 недели воздействия соединений в культуре (дополнительный рис.S7F). Поэтому мы исследовали, как c-MYC восстанавливает пролиферацию, выполнив анализ секвенирования РНК (RNA-seq) из цельного кишечника APC KRAS и APC KRAS MYC, обработанных с использованием или без eFT508 / рапамицина. Это выявило несколько изменений в количестве мРНК (68 измененных транскриптов) после экспрессии c-MYC в отсутствие лечения, но сравнение APC KRAS и APC KRAS MYC кишечника, обработанного комбинацией, выявило 335 измененных транскриптов (154 вверх, 181 вниз; дополнительный рис. S7G). Из этих транскриптов только девять были изменены экспрессией трансгена в отсутствие лекарственного лечения.REACTOME и анализ обогащения набора генов показал, что транскрипты, активируемые c-MYC, регулируют метаболические пути, такие как окислительное фосфорилирование и метаболизм арахидоновой кислоты (ссылки 47, 48; рис. 7E; дополнительный рис. S7H). Не было обогащения за изменения канонических c-MYC-модулированных транскриптов, таких как рибосомные составляющие или РНК-полимеразы. Повышение транскрипции специфических связанных с метаболизмом транскриптов было подтверждено с помощью количественной ПЦР из кишечной ткани, а также органоидов, обработанных eFT508 / рапамицином в течение одной недели (дополнительный рис.S7F и S7I). Интересно, что трансген c-MYC не изменяет экспрессию гена, чтобы напоминать необработанный контрольный APC KRAS кишечник, о чем свидетельствуют 865 транскриптов, экспрессируемых по-разному (573 вверх, 295 вниз) при сравнении этих двух условий (дополнительный рисунок S7G). Таким образом, трансген c-MYC специфически управляет программой транскрипции, направленной на метаболизм только после лечения eFT508 / рапамицином, что, вероятно, позволяет восстановить пролиферацию. Этот анализ даст представление о механизмах резистентности, управляемых c-MYC, и выявит окислительное фосфорилирование и метаболизм арахидоновой кислоты в качестве потенциальных мишеней.c-MYC регуляция метаболизма арахидоновой кислоты ранее наблюдалась на моделях Kras -мутантных мышей рака легких (49), а путь арахидоновой кислоты является протуморигенным в доклинических моделях колоректального рака, при этом ингибиторы, такие как целекоксиб, клинически эффективные профилактические методы лечения (50).
Учитывая эффективность воздействия на МНК в комбинации с рапамицином в модели APC KRAS, мы применили это лечение к более сложным моделям мышей колоректального рака, несущим дальнейшие мутации, распространенные при заболевании человека.Для этого мы использовали органоиды APC KRAS, также не содержащие Trp53 и Tgfbr1 , получившие название AKPT ( VillinCre ER Apc fl / fl Kras G12D / + Trp53 fl / fl Trgf fl ), и органоиды, выделенные из метастазов печени из модели автохтонного колоректального рака KPN ( VillinCre ER Kras G12D / + Trp53 fl / fl R26 N1ICD ; ссылка 51). В обоих случаях eFT508 / рапамицин значительно подавлял пролиферацию (дополнительный рис.S7J), расширяя эффективность лечебной комбинации за счет включения дополнительных мутаций рака.
Повышенная передача сигналов MNK и mTORC1 предсказывает плохую выживаемость
Затем мы исследовали переводимость двойного нацеливания на MNK и mTORC1 у пациентов с колоректальным раком. Таким образом, мы проанализировали выживаемость пациентов в отношении P-eIF4E и P-RPS6 и обработали органоиды пациентов с колоректальным раком ингибиторами MNK и mTORC1. Ни P-eIF4E, ни P-RPS6 по отдельности не связаны с выживаемостью в полной когорте проанализированных пациентов с колоректальным раком (дополнительный рис.S7K). Однако при стратификации с помощью GMS уровни P-RPS6 были значительно связаны с плохой специфической для рака выживаемостью у пациентов с GMS1 (опухоли с низким воспалительным инфильтратом и низким процентом опухоль-строма; дополнительный рисунок S7K). Опухоли GMS1 очень эпителиальны, с наименьшей вероятностью будут реагировать на иммунотерапию и часто имеют несколько вариантов лечения. Доклинические модели мышей, которые мы использовали, присутствовали с высокоэпителиальными опухолями, хорошо коррелирующими с опухолями GMS1. P-RPS6 и P-eIF4E положительно коррелируют в этих клинических образцах, и наши данные на животных предсказывают синергию между двумя путями, способствующую пролиферации.Поэтому мы объединили два маркера в единый балл, сравнивая выживаемость пациентов с опухолями, демонстрирующими низкое фосфорилирование одного или обоих маркеров, с пациентами с высоким фосфорилированием обоих маркеров. У пациентов с GMS1 с высоким уровнем передачи сигналов MNK и mTORC1 прогноз был значительно хуже, чем у пациентов с низким уровнем экспрессии (рис. 7F). В самом деле, средняя выживаемость больных с раком у «обоих высоких» пациентов была более чем на 3,5 года короче, чем у «одного или обоих пациентов с низким уровнем». Эти «оба высокие» пациенты с GMS1 с плохой выживаемостью составляли каждого пятого пациента в TMA.Во всей когорте передача сигналов через оба пути не имела значительного преимущества, что свидетельствует о специфичности к опухолям GMS1 (дополнительная фигура S7K).
Наконец, мы применили ингибиторы MNK и mTORC1 к двум полученным от пациентов линиям органоидов с делецией APC и KRAS G12D мутаций (52). Эти органоиды точно воспроизводят реакцию пациента на лечение, обеспечивая отличную систему для открытия клинически переводимых терапевтических средств.Обе линии органоидов были невосприимчивы к лечению рапамицином, но ответили на комбинацию eFT508 / рапамицин 20% снижением активности Cell-Titer Blue через 24 часа (дополнительный рисунок S7L). eFT508 (30 нмоль / л) полностью подавлял P-eIF4E в этих органоидах человека, и комбинированная обработка приводила к снижению экспрессии c-MYC в одной из линий (дополнительный рисунок S7M). В соответствии с эффектом eFT508 на эти органоиды человека, лечение мерестинибом снижает пролиферацию в комбинации с рапамицином в обеих линиях (дополнительный рис.S7L). В целом успешное нацеливание на mTORC1 и путь MNK / eIF4E, использованное в наших доклинических моделях мышей, имеет отличный потенциал для использования в клинике, где мы видим, что усиление передачи сигналов mTORC1 и MNK коррелирует с плохой выживаемостью в популяции пациентов с колоректальным раком.
Обсуждение
Хотя мутанты G12C теперь могут быть нацелены, они встречаются только в 3% всех случаев колоректального рака (4), а это означает, что большинство KRAS -мутантных колоректальных раков не имеют эффективных клинических вариантов.Чтобы решить эту проблему, мы стремились нацелить синтез белка. Мы показываем, что активация KRAS в кишечнике мышей увеличивает синтез белка, обеспечивая устойчивость к рапамицину, что согласуется с устойчивостью к рапалогам в клинике (7–9). Мы показываем, что механизм действия рапамицина изменяется мутацией Kras , при этом удлинение трансляции не затрагивается, но инициация трансляции подавляется. Наши данные также показывают несвязанность синтеза и пролиферации белка с пониженной трансляцией в нашей модели APC KRAS в ответ на рапамицин, но не влияет на пролиферацию.Мы продолжаем показывать, что сигнальные пути, регулирующие механизм трансляции, являются жизнеспособными мишенями в KRAS -мутантном колоректальном раке путем ингибирования P-eIF4E в комбинации с рапамицином, что снижает пролиферацию в GEMM множественного колоректального рака. Эта комбинация подавляла экспрессию c-MYC, что ставило под угрозу пролиферацию опухоли, с экспрессией человеческого трансгена C-MYC , полностью сводя на нет пользу лечения. Экспрессия c-MYC трансгена находится в физиологическом диапазоне с незначительной разницей или отсутствием разницы в уровнях c-MYC между аденомами мышей с опухолевыми моделями APC KRAS и APC KRAS MYC.Мы объясняем это тем, что чрезмерная экспрессия c-MYC сдерживается для защиты от апоптоза, индуцированного MYC (53).
Внутри клеток APC KRAS экспрессия c-MYC поддерживается за счет высоких скоростей синтеза белка и P-eIF4E. Снижение синтеза белка с помощью рапамицина переносится опухолями из-за того, что P-eIF4E поддерживает уровни c-MYC. Точно так же нацеливание только на P-eIF4E не подавляет экспрессию c-MYC, вероятно, из-за продолжающейся способности к трансляции. Однако снижение синтеза белка и ингибирование P-eIF4E приводит к снижению экспрессии c-MYC и меньшей пролиферации.Ингибирование либо mTORC1, либо MNKs ранее участвовало в регуляции трансляции C-MYC (23, 54-56), и ранее было показано, что нацеливание на комбинацию подавляет IRES-зависимую трансляцию c-MYC (57). Примечательно, что используемый здесь трансген C-MYC человека не имеет 5′-UTR и, следовательно, IRES, что дает возможность для будущих исследований механизма, лежащего в основе eFT508 / рапамицин-зависимого подавления c-MYC. Экспрессия белка c-MYC сдерживается комплексом eIF2B в тех же моделях мышей с колоректальным раком, которые используются здесь, при этом истощение eIF2B5 приводит к увеличению экспрессии c-MYC и MYC-зависимому апоптозу (58).Это подчеркивает важность трансляции в определении экспрессии c-MYC при колоректальном раке и, вместе с нашей работой, позиционирует модуляцию c-MYC ниже нацеленности на синтез белка.
Котаргетинг MNK и mTORC1 или других киназ снижает пролиферацию или образование колоний в клеточных линиях от различных видов рака (20, 38, 39, 59–63). Мы предоставляем первые доказательства эффективности этой комбинации препаратов на доклинической модели in vivo . Механизм действия этой комбинации препаратов не был полностью выяснен в предыдущих исследованиях; данные в некоторых из них согласуются с нашими наблюдениями, что нацеливание на MNKs и mTORC1 сходится при инициации трансляции и согласуется с делецией или ингибированием MNK, подавляющим глобальный синтез белка (20, 38, 39).eFT508 (томивосертиб) находится в фазе II клинических испытаний в сочетании с ингибиторами контрольных точек против солидных опухолей, включая колоректальный рак, и с паклитакселом при раке груди. Показанная здесь эффективность eFT508 / рапамицина для подавления пролиферации и увеличения выживаемости мышей более чем в 20 раз является обнадеживающей в качестве потенциальной терапии для пациентов с мутантным колоректальным раком KRAS . Комбинация также эффективна для органоидов колоректального рака, полученных от пациентов, которые точно воспроизводят лекарственные реакции.Мы также идентифицировали популяцию пациентов с колоректальным раком (> 45%) с одновременной передачей сигналов mTORC1 и MNK и наблюдали, что пациенты с высокоэпителиальными опухолями чувствуют себя значительно хуже, когда эти пути активированы.
В наших моделях передача сигналов через MNKs управляет пролиферацией раковых клеток присущим клеткам образом. Ингибирование MNK также подавляет активацию нейтрофилов в ответ на множественные стимулы (64) и ингибирует продукцию TNFα в Т-клетках (65), а в модели рака молочной железы мыши eIF4E KI имеют меньше прометастатических нейтрофилов и метастазов (66).P-eIF4E также играет внутреннюю роль раковых клеток в стимулировании экспрессии PD-L1 в мутантной модели гепатоцеллюлярной карциномы Kras (24). Нацеливание на P-eIF4E с использованием eFT508 снизило уровни PD-L1 и ослабило рост первичной опухоли и метастазирование. Возможно, будет дополнительная польза от нацеливания как на пролиферацию опухолевых клеток, так и на активность иммунных клеток через P-eIF4E.
Мы видим высокий уровень P-eIF4E у ~ 70% пациентов, что согласуется с предыдущим исследованием, в котором P-eIF4E был обнаружен в ~ 60% образцов колоректального рака человека (67).Как и при колоректальном раке, P-eIF4E увеличивается при плоскоклеточном раке головы и шеи (HNSCC) и раке легких (67). Аденокарциномы легких обнаруживают мутации в KRAS , а HNSCC обнаруживают мутации в своем паралоге HRAS (68, 69), причем экспрессия MYC также важна для обоих типов опухолей. Используемый здесь успешный терапевтический режим может быть полезным для пациентов с мутантным раком RAS , который проявляется с повышенными уровнями P-eIF4E и зависит от экспрессии MYC.Эта работа открывает возможность воздействовать на онкогенную передачу сигналов RAS во многих типах опухолей, воздействуя на его нижележащие регуляторы трансляции мРНК.
Методы
Доступность материалов
Все линии мышей, где это возможно, доступны по запросу, но могут подлежать оплате и / или соглашению о передаче материалов.
Клинические исследования
Это исследование было одобрено Комитетом по этике исследований Западной Шотландии (16 / WS / 0207), а информация о пациентах хранится в Большом Глазго и Клайд Сейф Хейвен (12 / WS / 0142).В ТМА использовалось четыре стержня диаметром 0,6 мм на каждого пациента. Для определения стадии опухоли использовали 5-е издание системы AJCC / UICC-TNM. GMS был построен путем комбинирования процентного содержания опухоли и стромы и баллов Клинтрупа-Мякинена, как описано ранее (11). В исследование были включены 102 пациента I – III стадии, перенесших лечебную операцию по удалению опухоли колоректального рака в Глазго Royal Infirmary, Western Infirmary или Stobhill Hospitals (Глазго, Великобритания) в период с 1997 по 2007 год. Пациенты, которые умерли в течение 30 дней после операции или прошли неоадъювантную терапию, были исключены.Пациенты наблюдались в течение как минимум пяти лет после резекции, при этом 49 случаев смерти от рака. Все имели действительные баллы для P-RPS6 S240 / 4 и 191 для P-eIF4E S209. ТМА окрашивали методом дуплексной хромогенной ИГХ на автоокрашивании Ventana Discovery Ultra (Roche) с использованием реагентов и систем обнаружения Ventana и следующих антител: P-RPS6 S240 / 4 [Cell Signaling Technology (CST) 5364], P-eIF4E S209 (Abcam ab76256) и цитокератин (Leica AE1 / AE3-L-CE). Слайды были отображены на Hamamatsu Nanozoomer XR и проанализированы с помощью Visiopharm версии 2019.02.2.6239. Чтобы очертить области опухоли, было разработано приложение контролируемой кластеризации K-средних. Приложение «Лес решений» было разработано для обнаружения клеток в опухолях, а окрашивание цитоплазмы было разделено по интенсивности для расчета H-балла. Кривые ROC были построены в MEDCalC, и индекс Юдена J был использован для определения границы между высоким и низким окрашиванием для обоих антител.
Исследования на мышах
Эксперименты проводились по лицензии Министерства внутренних дел Великобритании (60/4183 и 70/8646). Генотипирование мышей осуществляли с помощью Transnetyx.Все колонии были инбредными C57BL / 6J (поколение ≥7), за исключением колоний с аллелями eIF4E S209A или Rosa26-lsl-MYC , которые были аутбредными. Мышей содержали в обычных клетках с 12-часовым циклом свет / темнота и доступом к диете и воде ad libitum . Эксперименты проводились на самцах и самках в возрасте от 6 до 12 недель. Исследователи не были ослеплены. Размеры образцов указаны на рисунках или в подписях. Villin CreER -опосредованная рекомбинация была индуцирована внутрибрюшинным (т.е.p.) инъекция тамоксифена, растворенного в кукурузном масле, в дозе 80 мг / кг (70). Для ортотопической индукции 70 мкл 100 нмоль / л 4-ОН тамоксифена вводили в подслизистую оболочку толстой кишки с помощью светодиодного эндоскопа TelePack VetX (Karl Storz; ссылка 36). Эксперименты на опухолевых моделях были инициированы однократной дозой тамоксифена в день 0, за мышами наблюдали до появления признаков кишечного заболевания — потери веса, побледнения ног от анемии и сутулости. Эксперименты по лечению больных начинали, когда мыши теряли> 3% от своего стабильного веса.Мышей последовательно набирали на лечение носителем или лекарственным препаратом, чтобы гарантировать отсутствие систематической ошибки и равные размеры когорт. Опухоли оценивали макроскопически после фиксации открытой ткани кишечника. Для краткосрочных экспериментов мышей дикого типа для Kras индуцировали в последовательные дни (0 и 1), а затем отбирали пробы на 4-й день. Мышей, несущих аллель Kras G12D , индуцировали однократной инъекцией тамоксифена и образец взят на 3-й день после индукции. Животные были случайным образом разделены на группы, принимавшие наркотики, что обеспечивало равное гендерное разделение между когортами.Рапамицин (LC Laboratories R-5000) вводили ежедневно внутрибрюшинно. при 10 мг / кг (5). Мерестиниб (Axon MedChem 2553) суспендировали в 10% гуммиарабике и вводили в дозе 12 мг / кг ежедневно через желудочный зонд (OG; ссылка 23). eFT508 (производимый на собственном предприятии) суспендировали в 1% метилцеллюлозе, 0,15% Tween-80 и вводили OG дважды в день в дозе 1 мг / кг. Реагент для мечения пролиферации клеток BrdU (250 мкл; Amersham Bioscience RPN201) вводили i.p. 2 часа до отбора проб.
Гистология и IHC
Ткань фиксировали в формалине и заливали парафином.ИГХ проводили, как описано ранее (5), с использованием следующих антител: P-eIF4E S209 (Millipore 04-1058), RPS6 P-S240 / 4 (CST 5364), P-4E-BP1 (CST 2855), P- eEF2 T56 (Novus Biologicals NB100-92518), BrdU (BD Biosciences 347580), P-eIF4B S422 (Abcam ab59300), c-MYC (Santa Cruz Biotechnology Sc-764) и RFP (Tebu Bio 600–401–379). Было проанализировано не менее трех биологических повторов и отображены репрезентативные изображения. Для оценки BrdU в краткосрочных модельных экспериментах ткань фиксировали в смеси метанол: холоформ: уксусная кислота в соотношении 4: 2: 1, переносили в формалин и заливали парафином.Для оценки BrdU в модели опухоли фиксированную формалином залитую парафином ткань анализировали с использованием программного обеспечения Halo, обученного количественному определению BrdU-положительных ядер в эпителиальных опухолевых клетках в процентах от всех эпителиальных опухолевых клеток. Анализ BaseScope проводился в соответствии с инструкциями производителя (ACD) с использованием пользовательских зондов для мышиных Mnk1 оснований 658–789 и Mnk2 оснований 727–888.
Культура клеток
Культуры органоидов выделяли и поддерживали в Matrigel (Corning 356231).В среду Advanced DMEM / F12 (Life Technologies 12634-028) добавляли 5 ммоль / л HEPES (Life Technologies 15630-080), 2 ммоль / л l-глутамин (Life Technologies 25030-024), 100 Ед / мл пенициллин / стрептомицин. (Life Technologies 1540-122), 1 × N2 (Invitrogen 17502-048), 1 × B27 (Invitrogen 12587-010), 100 нг / мл ноггин (PeproTech 250-38) и 50 нг / мл EGF (PeproTech AF- 100–15). Органоиды, полученные от пациентов, выращивали в той же среде с добавлением 500 нг / мл R-спондина (R&D Systems 3474-RS), 10 нмоль / л гастрина (Sigma-Aldrich G9145), 100 нг / мл Wnt-3A (R&D Systems 5036). -WN), 10 мкмоль / л Y-27632 (Sigma-Aldrich Y0503), 0.5 мкмоль / л A83-01 (PeproTech 2939), 5 мкмоль / л SB202190 (Sigma-Aldrich S7067), 4 ммоль / л никотинамида (Sigma-Aldrich N0636), 10 нг / мл FGF basic (PeproTech 100-18B), 10 нг / мл FGF10 (PeproTech 100-26A), 1 мкмоль / л простагландина E2 (PeproTech 2296). Для анализа пролиферации клеток клетки помещали в 96-луночные планшеты, установленные на 2–3 дня перед использованием CellTiter-Blue для измерения метаболической емкости в качестве заменителя количества клеток (Promega G8080). Лекарства [5-ФУ (Mayne Pharma Plc PL04515 / 0088), оксалиплатин (Acoord Healthcare Ltd PL20075 / 0112) и рокагламид (Sigma-Aldrich SML0656)] вводили в течение 24 часов до добавления CellTiter-Blue для последних 2– 6 часов.Изменения метаболической емкости нормализовали для клеток, обработанных носителем. Все эксперименты с органоидами проводили до пассажа 20. Клетки HCT116 культивировали в среде DMEM с добавлением 2 ммоль / л 1-глутамина и 10% FBS (Life Technologies). Тестирование линий на Mycoplasma проводили с помощью MycoAlert (Lonza LT07-218).
РНК Иммунопреципитация
Органоиды собирали в ледяном PBS и матригеле и промывали ледяным PBS. Лизис проводили в буфере RIP [20 ммоль / л Tris-HCL pH 7.5, 200 ммоль / л NaCl, 5 ммоль / л MgCl 2 , 0,5% Triton X-100, смесь ингибиторов протеаз (Roche 11836153001), 800U ингибитор РНКазы RiboLock (Thermo Fisher Scientific [TFS] EO0381), 10% BSA, 0,5 мкмоль / л DTT, 5 мкмоль / л NaF] с 8-кратными проходами через иглу 21-го размера. Лизаты центрифугировали, определяли концентрацию белка в супернатанте и использовали равную концентрацию для каждого RIP. Кроме того, были собраны образцы для экстракции РНК и белка. Антитела, контроль IgG (CST 2729), eIF4E (TFS MA1089) и P-eIF4E S209 (Abcam ab76256), инкубировали с Dynabeads Protein G (TFS 10004D) в течение 2.5 часов. Аффинную очистку проводили в течение 30 минут при 4 ° C с постоянным вращением перед промывкой 3х буфером RIP. Полученную элюцию разделяли для экстракции белка или РНК с помощью TRIzol (TFS 10296010). Целостность РНК оценивали с помощью ленты RNA ScreenTape (Agilent Technologies 5067-5576) на системе 4200 TapeStation System. Конструкции FLAG-eIF4E были созданы путем амплификации человеческого eIF4E из плазмиды Addgene № 17343 с использованием 5′-CTGGAGATCTGATGGCGACTGTCGAACCGGAAACCAC-3 ‘и 5′-GCGTGGATCCTTAAACAACAACAAACCTATTTTTAGTGGTII’ 900×13 и 1,900-дигеста с помощью BAGTGGT-9001 и 900x1Digest-3gGAGI-9001 и 900×1 Вектор CMV (Sigma-Aldrich E7533).Мутация S209A была достигнута путем сайт-направленного мутагенеза с использованием QuikChange Lightning (Agilent 210518) и следующих праймеров: 5′-CGCAGACAGCTACTAAGAGCGGCGCCACCACTAAAAATAG-3 ‘и 5’-CTATTTTTAGTGGTGCGCCGCTCGTCTAGTAGTAG. Плазмиды трансфицировали в клетки HCT116 с использованием липофектамина 2000 (TFS 11668019) в соответствии с инструкциями производителя. Через 24 часа после трансфекции 30 миллионов клеток на каждое условие собирали трипсинизацией. Его лизировали в буфере RIP, а затем очищенные лизаты смешивали с предварительно промытыми шариками, конъюгированными с антителом против FLAG (Merck M8823), в течение 30 минут при 4 ° C, промывали, а затем элюировали РНК в TRIzol.Образцы с параллельным вводом были сохранены, и РНК также очищали с помощью TRIzol. Контрольная плазмида p3xFLAG-BAP была предоставлена Sigma-Aldrich.
qPCR
РНК
была очищена из ткани с помощью RNeasy (Qiagen 74104) после разрушения в пробирках CK14 Precellys с расщеплением ДНКазой на колонке. Очищенную РНК подвергали обратной транскрипции (верхний индекс III) с использованием случайных гексамеров и количественной ПЦР, используя следующие праймеры для мышиных транскриптов. Mnk1 : прямой (Fd) 5′-CCATCGTGGATTCTGACAAGAG-3 ‘, обратный (Rv) 5′-GAACACTCGACTTCGACTGTG-3’; Mnk2 : Fd 5′-TCGGGCTACTGACAGCTTCT-3 ‘, Rv 5′-GACACAGGTCTGCACACGAG-3’; C-myc : Fd 5′-CCCAAATCCTGTACCTCGTC-3 ‘, Rv 5′-TTGCCTCTTCTCCACAGACA-3’; ActB : Fd 5′-GTGACGTTGACATCCGTAAAGA-3 ‘, Rv 5′-GCCGGACTCATCGTACTCC-3’; Циклин D1 : Fd 5′-GAGAAGTTGTGCATCTACACTG-3 ‘, Rv 5′-AAATGAACTTCACATCTGTGGC-3’; Циклин E : Fd 5′-GTGGCTCCGACCTTTCAGTC-3 ‘, Rv 5′-CACAGTCTTGTCAATCTTGGCA-3’; Mcl1 : Fd 5′-GACGACCTATACCGCCAGTC-3 ‘, Rv 5′-AGAGGCTTCGAGTCCTTGGA-3’; Snail1 : Fd 5′-CACACGCTGCCTTGTGTCT-3 ‘, Rv 5′-GGTCAGCAAAAGCACGGTT-3’; Runx2 : Fd 5′-AGAGTCAGATTACAGATCCCAGG-3 ‘, Rv 5′-TGGCTCTTCTTACTGAGAGAGG-3’; Gapdh : Fd 5′-TCCACTGGCGTCTTCACC-3 ‘, Rv 5′-GGCAGAGATGATGACCCTT-3’; Enpp7: Fd 5′-GACAGCATAAGCTACTCCTCGT-3 ‘, Rv 5′-GACAGCATAAGCTACTCCTCGT-3’; Slc36a1 : Fd 5′-GACTACAGTTCCACAGACGTG-3 ‘, Rv 5′-CCATGTCATGCTACTGCTCTCT-3’; Car4 : Fd 5′-TACGTGGCCCCCTCTACTG-3 ‘, Rv 5′-GCTGATTCTCCTTACAGGCTCC-3’.Для транскриптов человека использовали следующие праймеры: c-MYC : Fd 5′-TACAACACCCGAGCAAGGAC-3 ‘, Rv 5′-GAGGCTGCTGGTTTTCCACT-3’; Циклин D1 : Fd 5′-GAGAAGTTGTGCATCTACACTG-3 ‘, Rv 5′-AAATGAACTTCACATCTGTGGC-3’; Циклин E: Fd 5′-ACTCAACGTGCAAGCCTCG-3 ‘, Rv 5′-GCTCAAGAAAGTGCTGATCCC-3’; MCL1: Fd 5′-GCTCAAGAAAGTGCTGATCCC-3 ‘, Rv 5′-TAGCCACAAAGGCACCAAAAG-3’; ACTB : 5′-GCCAACAGAGAGAAGATGAC-3 ‘, Rv 5′-CGCAAGATTCCATACCCAGG-3’; TUBA1B : Fd 5′-AGCATCCAGTTTGTGGATTGGTGC-3 ‘, Rv 5′-CAAAGGCACGCTTGGCATACATCA-3’; GAPDH : Fd 5′-CTATAAATTGAGCCCGCAGCC-3 ‘Rv, 5′-ACCAAATCCGTTGACTCCGA-3’.
RNA-seq
Библиотеки для генерации кластеров и секвенирования ДНК были подготовлены в соответствии с ранее описанным методом (71) с использованием набора для подготовки образцов TruSeq RNA v2 (Illumina), затем запускались на Illumina NextSeq с использованием набора High Output 75 циклов ( 2 × 36 циклов, парные чтения, одиночный индекс). Качество необработанной последовательности оценивалось с помощью FastQC версии 0.11.8, затем последовательности были обрезаны для удаления последовательностей адаптеров и низкокачественных базовых вызовов, определенных как те, у которых показатель Phred менее 20, с использованием Trim Galore версии 0.6.4. Обрезанные последовательности выравнивали с построением генома мыши GRCm38.98 с использованием HISAT2 версии 2.1.0, и исходные количества на ген определяли с использованием FeatureCounts версии 1.6.4. Анализ дифференциальной экспрессии выполнялся с использованием пакета R DESeq2 версии 1.22.2, затем анализ пути проводился для значительно измененных генов (определяемых как FDR <0,05) с использованием пакета R ReactomePA версии 1.26.0. Данные, обсуждаемые в этой публикации, депонированы в омнибусе экспрессии генов NCBI, доступном через регистрационный номер серии GEO GSE159432.
Вестерн-блоттинг
Образцы лизировали [10 ммоль / л Трис (pH 7,5), 50 ммоль / л NaCl, 0,5% NP40, 0,5% SDS с добавлением коктейля ингибиторов протеазы, PhosSTOP (Roche 047001) и бензоназы (Sigma-Aldrich). E1014)] на льду, а количество белка оценивали с помощью анализа BCA (TFS 23225). Двадцать микрограммов белка денатурировали в SDS-красителе и затем разделяли 4–12% SDS-PAGE (Invitrogen NP0336BOX). Белок переносили в нитроцеллюлозу и подвергали иммуноблоттингу в течение ночи при 4 ° C с использованием следующих антител: eIF4E (CST 2067), P-eIF4E S209 (CST 9741), RPS6 (CST 2317), RPS6 P-S240 / 4 (CST 5364), 4E -BP1 (CST 9644), eEF2 (CST 2332), P-eEF2 T56 (CST 2331), eIF4B (CST 3592), P-eIF4B S422 (CST 3591), eIF4G1 (CST 2498), P-eIF4G1 S1108 (CST 2441) ), FLAG (Abcam ab1162), винкулин (ab18058 Abcam) и β-актин (Sigma-Aldrich A2228).Вторичные антитела, конъюгированные с пероксидазой хрена (Dako P0447, P0448), инкубировали в течение 1 часа при комнатной температуре, а затем мембраны подвергали рентгенографии в электрохемилюминесценции (TFS 32106). Количественную оценку выполняли с использованием ImageJ (NIH, Bethesda, MD).
Градиенты плотности сахарозы
Органоиды добавляли средой в течение шести часов, а затем в среду добавляли 200 мкг / мл циклогексимида (Sigma-Aldrich C7692) в течение трех минут перед сбором на льду.Фракции крипт выделяли путем экстракции эпителия из 10 см проксимального отдела тонкой кишки. Ворсинки удаляли, инкубируя промытые PBS линейно открытый кишечник в среде RPMI (TFS 21875059) с добавлением 10 ммоль / л EDTA и 200 мкг / мл циклогексимида в течение 7 минут при 37 ° C при регулярном перемешивании. Затем крипты выделяли переносом ткани в ледяной PBS, содержащий 10 ммоль / л EDTA и 200 мкг / мл циклогексимида, на 7 минут. Затем оставшуюся ткань выбросили. Образцы лизировали [300 ммоль / л NaCl, 15 ммоль / л MgCl 2 , 15 ммоль / л Трис pH 7.5, 100 мкг / мл циклогексимида, 0,1% тритона X-100, 2 ммоль / л DTT и 5 ед / мл SUPERaseIn (TFS AM2696)] на льду и постъядерные экстракты, наслоенные на 10% –50% вес / объем сахарозы градиенты, содержащие тот же буфер без Triton X-100, DTT или SUPERaseIn. Градиенты вращали при 255000 rcf в течение 2 часов при 4 ° C под вакуумом в роторе SW40Ti. После центрифугирования образцы разделяли с помощью считывающего устройства с оптической плотностью 254 нм (ISCO). Отношения полисомов к субполисомам (P: S) были рассчитаны с использованием метода трапеций с указанием границ вручную.Для анализа стекания харрингтонина культуры готовили в двух экземплярах, одну предварительно обрабатывали 2 мкг / мл харрингтонина (Santa Cruz Biotechnology sc-204771) в течение 5 минут (300 секунд) перед добавлением циклогексимида, а затем обе обрабатывали, как указано выше. Скорость стекания рассчитывалась как соотношение P: S без харрингтонина по сравнению с соотношением P: S с харрингтонином.
35 Мечение S-метионином
Органоиды анализировали во время оптимальной фазы роста, через 3 дня после разделения и через 6 часов после смены среды. 35 В S-метионин (PerkinElmer NEG772002MC) добавляли 30 мкКи / мл в течение 30 минут перед сбором и лизировали с использованием буфера для вестерн-блоттинга. Белок осаждали в 12,5% (мас. / Об.) Трихлоруксусной кислоте на бумаге из стекловолокна (Whatmann 1827-024) с использованием вакуумного коллектора и промывали 70% этанолом и ацетоном. Сцинтилляцию считывали на счетчике Wallac MicroBeta TriLux 1450 с использованием Ecoscint (SLS Ltd LS271) и нормализовали по общему содержанию белка, определенному с помощью анализа BCA. Скорость синтеза белка выражали как сцинтилляцию над содержанием белка (CPM / мкг / мл белка) относительно контрольного образца, установленного на 1.
Статистический анализ
Статистический анализ и номера n выделены на каждом рисунке или в легенде. Во всех случаях значения P , равные или менее 0,05, считались статистически значимыми.
Стоит ли у вас возникнуть соблазн купить публичную компанию с ограниченной ответственностью Mallinckrodt (NYSE: MNK) из-за ее коэффициента PE?
Этот анализ предназначен для ознакомления с важными ранними концепциями людям, которые начинают инвестировать и хотят начать изучать основные концепции фундаментального анализа на практических примерах сегодняшнего рынка.
Публичная компания с ограниченной ответственностью Mallinckrodt (NYSE: MNK) в настоящее время торгуется с скользящим коэффициентом P / E 1,8, что ниже среднего по отрасли 28,2. Хотя это делает MNK хорошей акцией для покупки, вы можете передумать после того, как я объясню предположения, лежащие в основе отношения P / E. Сегодня я проанализирую соотношение P / E и выделю то, чего вам следует остерегаться при использовании отношения P / E.
Ознакомьтесь с нашим последним анализом для Mallinckrodt
Демистификация отношения P / E
NYSE: MNK PE PEG Gauge 31 августа 18
P / E — популярный коэффициент, используемый для относительной оценки.Он сравнивает цену акции на акцию с прибылью на акцию. Более интуитивно понятный способ понять коэффициент P / E — это подумать о том, сколько инвесторы платят за каждый доллар прибыли компании.
Формула
Соотношение цены и прибыли = Цена на акцию ÷ Прибыль на акцию
Расчет P / E для MNK
Цена на акцию = 34,09 доллара
Прибыль на акцию = 19,27 доллара
∴ Цена — Цена = 34 доллара США.09 ÷ 19,27 доллара США = 1,8x
Коэффициент P / E не является показателем, который вы рассматриваете изолированно, и становится полезным только при его сравнении с другими аналогичными компаниями. В идеале мы хотим сравнить коэффициент P / E акций со средним показателем компаний, которые имеют такие же характеристики, как MNK, такие как размер и страна операций. Быстрый метод создания группы сверстников — использовать компании из одной отрасли, чем я и займусь. Поскольку аналогичные компании технически должны иметь схожие коэффициенты P / E, мы можем очень быстро прийти к некоторым выводам об акциях, если коэффициенты различаются.
Поскольку коэффициент P / E MNK составляет 1,8 ниже, чем у аналогичных компаний по отрасли (28,2), это означает, что инвесторы платят меньше за каждый доллар прибыли MNK. Этот коэффициент является средним для прибыльных компаний 24 фармацевтических компаний в США, включая Osteologix Holdings, Biostar Pharmaceuticals и Prestige Consumer Healthcare. Вы можете думать об этом так: рынок предполагает, что MNK — более слабый бизнес, чем средняя сопоставимая компания.
Допущения, о которых необходимо знать
Прежде чем делать поспешные выводы, важно понять, что наши предположения основаны на двух важных утверждениях.Во-первых, наши «похожие компании» фактически похожи на МНК. Если компании не похожи, разница в P / E может быть результатом других факторов. Например, если вы случайно сравнили компании с более высокими темпами роста с MNK, то P / E MNK, естественно, будет ниже, поскольку инвесторы вознаградят более высокие темпы роста компаний-аналогов более высокой ценой. В качестве альтернативы, если вы непреднамеренно сравните менее рискованные фирмы с MNK, P / E MNK снова будет ниже, поскольку инвесторы будут вознаграждать меньший риск его коллег более высокой ценой.Второе предположение, которое должно выполняться, заключается в том, что акции, с которыми мы сравниваем MNK, справедливо оценены рынком. Если это предположение не соответствует действительности, более низкий коэффициент P / E MNK может быть вызван тем, что компании из нашей группы компаний-аналогов переоценены рынком.
NYSE: MNK Future Profit 31 августа 18
Что это означает для вас:
Если ваше личное исследование акций подтверждает, что вам говорит соотношение P / E, возможно, сейчас самое время добавить больше MNK в ваш портфель . Но имейте в виду, что полезность относительной оценки зависит от того, готовы ли вы делать предположения, о которых я упоминал выше.Помните, что основывать свое инвестиционное решение только на одном показателе явно недостаточно. Есть много вещей, которые я не учел в этой статье, и отношение PE очень одномерно. Если вы еще этого не сделали, я призываю вас завершить свое исследование, взглянув на следующее:
- Перспективы на будущее : Что хорошо информированные отраслевые аналитики прогнозируют в отношении будущего роста MNK? Взгляните на наш бесплатный исследовательский отчет о консенсусе аналитиков в отношении прогнозов MNK.
- Прошлый послужной список : Показывает ли MNK стабильно хорошие результаты независимо от взлетов и падений на рынке? Более подробно ознакомьтесь с прошлым анализом производительности и взгляните на бесплатные визуальные представления истории MNK для большей ясности.
- Другие высокодоходные акции : Существуют ли другие акции, обеспечивающие более высокие перспективы с подтвержденным послужным списком? Ознакомьтесь с нашим бесплатным списком этих замечательных акций здесь.
Чтобы помочь читателям увидеть краткосрочную волатильность финансового рынка в прошлом, мы стремимся предоставить вам долгосрочный целенаправленный исследовательский анализ, основанный исключительно на фундаментальных данных.Обратите внимание, что в нашем анализе не учитываются последние объявления компаний, чувствительных к ценам.
Автор является независимым участником и на момент публикации не имел позиции в указанных акциях. В случае ошибок, требующих исправления, обращайтесь к редактору по адресу [email protected].
Promoted
При торговле Mallinckrodt или любыми другими инвестициями используйте платформу, которую многие считают воротами профессионалов на мировой рынок, Interactive Brokers.Вы получаете самую дешевую * торговлю акциями, опционами, фьючерсами, валютой, облигациями и фондами по всему миру с единого интегрированного счета.
Simply Wall St не имеет позиций ни в одной из упомянутых компаний. Эта статья носит общий характер.