
Содержание
Ударение в слове РЕКУ: как правильно
Содержание: ударение в слове “РЕКУ”
“РЕКУ” – это форма винительного падежа единственного числа неодушевленного существительного “река”.
В слове начальной формы ударение падает на второй слог:
река́
Давайте теперь разберемся с произношением формы винительного падежа.
Для начала разобьем данное слово на фонетические слоги:
ре-ку
В слове “реку” 2 слога, 4 буквы и 4 звука.
Так какой же слог будет ударным в данном случае?
В разговорной речи бытует два варианта произношения анализируемого слова:
- “ре́ку”, где ударение падает на гласную букву “е” первого слога,
- “реку́“, где ударение падает на гласную букву “у” второго слога.
Как правильно – “рЕку” или “рекУ”?
Согласно орфоэпической норме русского языка ударение в слове “реку” допускается ставить как на первый слог, так и на второй.
ре́ку
реку́
Понаблюдаем за ударением в разных падежных формах единственного числа:
- и. п. кто? (что?) река́
- р. п. кого? (чего?) реки́
- д. п. кому? (чему?) реке́
- в. п. кого? (что?) ре́ку́
- т. п. кем? (чем?) реко́й (реко́ю)
- п. п. о ком? (о чем?) реке́
Теперь понаблюдаем за ударением в разных падежных формах множественного числа:
- и. п. кто? (что?) ре́ки
- р. п. кого? (чего?) рек
- д. п. кому? (чему?) ре́кам
- в. п. кого? (что?) ре́ки
- т. п. кем? (чем?) ре́ками
- п. п. о ком? (о чем?) ре́ках
Как мы видим, во всех падежах единственного числа ударение падает на второй слог. Исключение составляет лишь форма винительного падежа, где ударение является вариативным.
Что касается падежей множественного числа, то ударение всегда падает на первый слог.
Примеры для закрепления:
- Они целый час не могли переправиться через ре́ку́.

- Детей нельзя отпускать на ре́ку́ без сопровождения взрослых.
- Они пошли на ре́ку́, чтобы покататься на самодельной лодке.
Подготовка к ЕГЭ по русскому языку и ГИА
Мы думаем, что каждый, кто сдаёт единый государственный экзамен, хочет получить за него максимальное количество баллов. С хорошими результатами будет легче поступить в любой вуз. Данный раздел поможет вам приблизиться к этой цели. Здесь есть всё необходимое для успешной подготовки. Также данный раздел нередко используется учащимися вузов и ссузов.
Проверить орфографию онлайн
Математика
- Часть A:
- Согласные звонкие и глухие
- Ударение в словах
- Паронимы. Лексическое значение слов
- Склонение имен существительных, падежи русского языка
- Деепричастный оборот, примеры
- Нормы согласования и управления
- Последовательная связь предложений в тексте
- Сочетание слов. ЕГЭ по русскому языку
- Грамматическая основа предложений
- Подчинительная, сочинительная, бессоюзная связь
- Правописание причастий, разряды местоимений, предлоги, частицы
- Лексическое значение слов
- Суффиксы.
 Приставки. Виды, примеры, правописание
Приставки. Виды, примеры, правописание - Правописание суффиксов прилагательных, Н, НН
- Проверочные слова, безударные гласные в корне
- Правописание приставок
- Правописание безударных личных окончаний глагола
- Правописание суффиксов глаголов
- Правописание не или ни
- Правописание предлогов
- Однородные члены предложения
- Знаки препинания при обособленных согласованных определениях
- Вводные слова в предложении
- Знаки препинания при однородных членах
- Знаки препинания в предложениях
- A26
- A27
- Действительные и страдательные причастия
- Микротема, основная мысль текста
- Типы речи: описание, повествование, рассуждение
- Синонимы к словам
- Часть B:
- Бессуффиксный способ словообразования
- Определение части речи
- Типы подчинительной связи
- Безличные, определенно-личные, односоставные предложения
- Обособленные приложения, обстоятельства и примеры
- СПП с придаточными
- Средства связи частей текста
- Что такое эпитет метафора, сравнение
- Часть C:
- Сочинение ЕГЭ по русскому языку
Обществознание
За последние несколько лет тема единого государственного экзамена стала особенно актуальной. Изначально эта программа вводилась как эксперимент и уже в первые месяцы тестирования зарекомендовала себя как объективную систему тестирования выпускников. Так что же все-таки представляет из себя этот ЕГЭ?
Изначально эта программа вводилась как эксперимент и уже в первые месяцы тестирования зарекомендовала себя как объективную систему тестирования выпускников. Так что же все-таки представляет из себя этот ЕГЭ?
Например, ЕГЭ по русскому языку состоит из трех частей (А, B, C). В первой части (A) 30 вопросов с одним вариантом ответа, а в части В, более сложной, чем А, всего 8 вопросов с написанием правильного ответа или выбором нескольких ответов. Каждому выпускнику одиннадцатых классов в обязательном порядке следует сдавать только 2 предмета: русский язык и математика, остальные по выбору. Допускаются к экзамену только ученики, имеющие оценки не ниже удовлетворительных, то есть без двоек в аттестате. Проверка работ производится другими преподавателями в другом районе, дабы исключить всякую возможность коррупции.
В школах многие учителя буквально наводят ужас на своих учеников, рассказывая о беспощадности ЕГЭ, в большинство ВУЗов принимают только с определенным количеством баллов, а различные организации твердят о ЕГЭ, чтобы привлечь к себе клиентов, желающих получить достойную подготовку к экзамену. Должен сказать, что квалифицированная подготовка дает свои, далеко не плохие, результаты. Но те, кто уже прошел через это «страшное» испытание, утверждают, что для учеников даже со средними оценками экзамен не должен показаться слишком уж сложным, по крайней мере невыполнимым. Нужно лишь приложить немного усилий, а именно выучить хотя бы самые важные правила, пройденные за весь учебный период, ведь если вы не ленились и хотя бы иногда открывали учебники, то что-то вы должны знать. Очень хорошо помогают различные книжки, предлагающие собственные примеры заданий, примеры их решений и дающие различные рекомендации по сдаче экзамена. Подобной литературой буквально завалены все книжные магазины, причем стоят они очень дешево. Для кого-то, естественно, и этого будет недостаточно. В таких случаях я бы рекомендовал обращаться к своим учителям, большинство из которых готовы помогать бесплатно. Я знаю, что во многих школах учителя предлагают организовывать собственные школьные подготовительные курсы за небольшую плату, а то и вовсе бесплатно.
Должен сказать, что квалифицированная подготовка дает свои, далеко не плохие, результаты. Но те, кто уже прошел через это «страшное» испытание, утверждают, что для учеников даже со средними оценками экзамен не должен показаться слишком уж сложным, по крайней мере невыполнимым. Нужно лишь приложить немного усилий, а именно выучить хотя бы самые важные правила, пройденные за весь учебный период, ведь если вы не ленились и хотя бы иногда открывали учебники, то что-то вы должны знать. Очень хорошо помогают различные книжки, предлагающие собственные примеры заданий, примеры их решений и дающие различные рекомендации по сдаче экзамена. Подобной литературой буквально завалены все книжные магазины, причем стоят они очень дешево. Для кого-то, естественно, и этого будет недостаточно. В таких случаях я бы рекомендовал обращаться к своим учителям, большинство из которых готовы помогать бесплатно. Я знаю, что во многих школах учителя предлагают организовывать собственные школьные подготовительные курсы за небольшую плату, а то и вовсе бесплатно.
Что же касается ГИА, то тут тоже ничего особо сложного нет, разница лишь в том, что задания в работах немного легче и сам экзамен не так важен как ЕГЭ, ведь ГИА проводится только среди девятых классов.
В заключение хотелось бы сказать, что сдать экзамен не так сложно, как пугают учителя, но нельзя преуменьшать важность и серьезность ЕГЭ, а также степень легкости экзамена, ведь, как ни крути, а на раз плюнуть никакие экзамены не даются: всё требует подготовки и старания.
Ударение в слове РОЖКИ: как правильно
Содержание: ударение в слове “РОЖКИ”
“РОЖКИ” – это неодушевленное существительное, которое употребляется в нескольких значениях:
- маленькие рога животного
- макаронные изделия
Теперь давайте разберемся с правильным произношением данного слова.
Для начала разобьем анализируемое слово на фонетические слоги:
ро-жки
В слове “рожки” 2 слога, 5 букв и 5 звуков.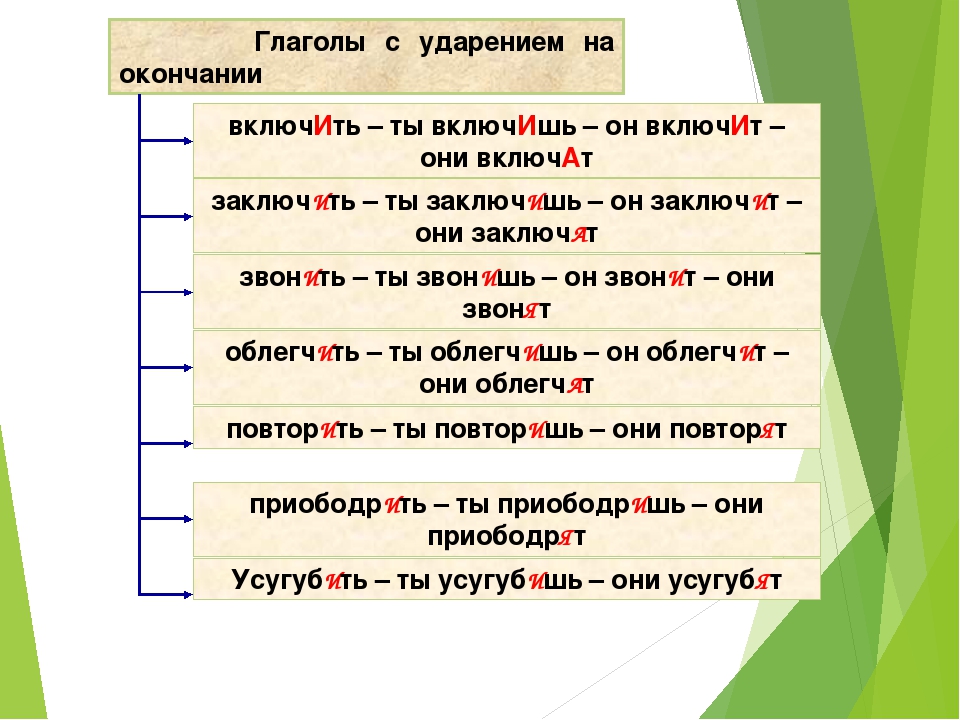
Так какой же слог будет ударным в данном случае?
В разговорной речи бытует два варианта произношения анализируемого слова:
- “ро́жки”, где ударение падает на гласную букву “о” первого слога,
- “рожки́“, где ударение падает на гласную букву “и” второго слога.
Как правильно – “рОжки” или “рожкИ”?
Согласно орфоэпической норме русского языка ударение в слове “рожки” ставится на первый или на второй слоги в зависимости от значения.
ро́жки
Ударение следует ставить на первый слог, если мы употребляем слово в значении “рога животного”.
Понаблюдаем за другими падежными формами множественного числа:
- и. п. кто? (что?) ро́жки
- р. п. кого? (чего?) ро́жек
- д. п. кому? (чему?) ро́жкам
- в. п. кого? (что?) ро́жки
- т. п. кем? (чем?) ро́жками
- п. п. о ком? (о чем?) ро́жках
Как мы видим, ударение и в косвенных падежах падает на первый слог.
рожки́
Ударение следует ставить на второй слог, если мы употребляем слово в значении “макаронные изделия”.
Понаблюдаем за ударением в разных падежных формах множественного числа:
- и. п. кто? (что?) рожки́
- р. п. кого? (чего?) рожко́в
- д. п. кому? (чему?) рожка́м
- в. п. кого? (что?) рожки́
- т. п. кем? (чем?) рожка́ми
- п. п. о ком? (о чем?) рожка́х
Как мы видим, во всех формах ударение падает только на второй слог.
Примеры для закрепления:
- У лосенка были очень маленькие и миленькие ро́жки.
- Рожки́ уже были остывшими, когда мама позвала за стол.
- Сегодня рожки́ были вкусными, как никогда.
Ударение в словах | ДИКТОРЫ.com
Особенности ударения в русском языке (продолжение)
Отдельного разговора требуют безударные слова. Служебные слова и частицы, как правило, не имеют на себе ударения в русском языке. Одни из них — односложные предлоги и союзы, бывают предударными словами, так называемыми проклитиками. Они примыкают в произношении к следующим за ними самостоятельным словам, имеющим ударение: на водЕ, при дорОге, от лЕса, по путИ. Другие — односложные частицы, являются энклитиками, то есть послеударными словами. Они примыкают в произношении к предшествующему слову, имеющему на себе удар.: пришел ктО-то, скажИ-ка, Я ведь обещал, тЫ же открыл дверь, придУт ли они. В этих сочетаниях частицы то, ка, ведь, же, ли становятся энклитиками.
Одни из них — односложные предлоги и союзы, бывают предударными словами, так называемыми проклитиками. Они примыкают в произношении к следующим за ними самостоятельным словам, имеющим ударение: на водЕ, при дорОге, от лЕса, по путИ. Другие — односложные частицы, являются энклитиками, то есть послеударными словами. Они примыкают в произношении к предшествующему слову, имеющему на себе удар.: пришел ктО-то, скажИ-ка, Я ведь обещал, тЫ же открыл дверь, придУт ли они. В этих сочетаниях частицы то, ка, ведь, же, ли становятся энклитиками.
Бывает, предлог принимает удар. на себя, тогда следующее за ним знаменательное слово оказывается безударным. Чаще всего перетягивают на себя удар. предлоги НА, ЗА, ПОД, ПО, ИЗ, БЕЗ.
НА — нА воду, нА гору, нА руку, нА ухо, нА зиму, нА год, нА дом, нА пол. Но такой перенос ударной гласной не всегда происходит. Мы говорим выйти на мОл (но не нА мол), взобраться на хОлм (но не нА холм), указать на двЕрь (но не нА дверь), сесть на мЕль (но не нА мель).
Перенос ударения на предлог, по нормам орфоэпии, возможен тогда, когда сочетание существительного с предлогом входит в состав устойчивого оборота или когда оно выступает в обстоятельственном значении и имеет наречный характер. В том же случае, когда важно выделить существительное как объект, на который направлено действие, и когда это существительное выступает в роли дополнения, удар. на предлог не переходит. Например:
поверить нА слово, но: обратить внимание на слОво «преобразование»;
спустить корабль нА воду, но: из-за солнечных бликов больно смотреть на вОду;
этот человек нечист нА руку, но: на рУку была наложена повязка;
взвалить груз нА плечи, но: он положил ладони ей на плЕчи;
сдвинуть шапку нА нос, но: жонглёр поставил на нОс картонный цилиндр;
старик туг нА ухо, но: мать посмотрела на Ухо мальчика.
Мы скажем взять грех нА душу. Это устойчивый оборот, и удар. в нем закреплено. Но нельзя говорить: столько-то продукции приходится нА душу населения. Мы говорим свалиться как снег нА голову. Это тоже фразеологизм, в котором ударение традиционно ставится на предлог. Но нельзя говорить: он бросил конфетти нА головы друзей.
Мы говорим свалиться как снег нА голову. Это тоже фразеологизм, в котором ударение традиционно ставится на предлог. Но нельзя говорить: он бросил конфетти нА головы друзей.
Часто ударение в русском языке переносится на предлог НА при сочетании его с числительными: нА два, нА три, нА пять, нА десять, нА сто, нА двое, нА трое. Но если рядом стоят два числительных со значением приблизительности, такого передвижения удар. не происходит: уехать на двА-трИ дня, на пЯть-шЕсть месяцев, на двОе-трОе суток. Произнесение нА два-три, нА три-пять — неверное. Предлог остается безударным и в том случае, когда два числительных соединены союзом ИЛИ: на двА или трИ дня, на пЯть или шЕсть месяцев, на двОе или трОе суток.
Не переносится ударение в русском языке на предлог и тогда, когда при первом числительном есть уточнение. Сравните: уехать нА два месяца — уехать на двА месяца и десять дней; командировка нА год — командировка на гОд и три месяца; встреча назначена нА три часа — встреча назначена на трИ часа тридцать минут.
Наконец, следует сказать, что иногда содержание текста подсказывает необходимость сохранить удар. на знаменательном слове, а не переносить его на предлог. В беседе о творчестве известного русского композитора речь шла о сюите на двЕ русские темы (а не нА две). Ведущий специально выделил слово две, чтобы обратить на это внимание аудитории.
ЗА — зА ногу, зА руку, зА зиму, зА душу, зА город, зА ночь, зА день, зА два, зА три, зА пять, зА семь, зА сорок.
Но и здесь те же ограничения:
спрятать руки зА спину — спрятаться за спИну матери;
схватить зА руки — схватить за рУки и нОги;
туда можно добраться зА два часа — за двА часа сорок минут.
ПО — пО лесу, пО полю, пО полу, пО носу, пО два, пО три, пО сто, пО двое, пО трое.
Не переносится удар. на предлог при сочетании его с числительными пять, шесть, семь, восемь, девять, десять… сорок и сложными числительными одиннадцать, двенадцать, пятьсот, шестьсот и т. д.: по пЯть часов, по шЕсть дней, по дЕвять рублей, по сОрок копеек.
д.: по пЯть часов, по шЕсть дней, по дЕвять рублей, по сОрок копеек.
Ударение не переносится и в описанных выше случаях:
по двЕ-трИ тарелки, по пЯть-шЕсть человек;
по двОе-трОе суток, по двА или трИ дня;
по двОе ши трОе суток, по трОе с половиной суток;
пО два центнера, но по двА и три десятых центнера.
ПОД — пОд ноги, пОд руки, пОд гору, пОд нос, пОд вечер.
ИЗ — Из дому, Из лесу, Из виду, Из носу.
БЕЗ — бЕз вести, бЕз году неделя, бЕз толку.
ОТ- час От часу, год От году, От роду.
Некоторые двусложные предлоги всегда бывают безударными. Таковы ИЗ-ЗА, ИЗ-ПОД и предлоги с беглым О: ПОДО, НАДО, ОБО, ОТО, ИЗО — из-за лЕса, из-под столА, подо мнОй, обо мнЕ, ото всЕх, изо всЕх.
Слабоударяемые слова — это те слова, которые хотя и имеют во фразе ударение, но более слабое, чем у самостоятельных слов. Такое ударение называют побочным. Слабоударяемыми словами бывают многие наречные предлоги, такие, как ПОСЛЕ, КРУГОМ, МИМО, ВОКРУГ, НАПРОТИВ, ПОПЕРЕК, ОКОЛО и другие. То, что на эти слова падает лишь ослабленное удар., ясно ощущается при сопоставлении с фразами, в которых слова эти, употребленные в качестве наречий, становятся носителями нормального удар.:
Слабоударяемыми словами бывают многие наречные предлоги, такие, как ПОСЛЕ, КРУГОМ, МИМО, ВОКРУГ, НАПРОТИВ, ПОПЕРЕК, ОКОЛО и другие. То, что на эти слова падает лишь ослабленное удар., ясно ощущается при сопоставлении с фразами, в которых слова эти, употребленные в качестве наречий, становятся носителями нормального удар.:
мать стояла сзади детЕй — река осталась сзАди;
поезд мчался мимо полЕй — машина проехала мИмо;
он помахал вслед автОбусу — люди что-то кричали вслЕд;
мы придём после чАса — мы придём к вам пОсле;
около дОма был родник — довольно ходить вокруг да Около.
Побочное ударение (или второстепенное) обозначается знаком «гравис», в отличие от основного удар., обозначаемого знаком «акут». Побочное удар. имеет обычно предлог СКВОЗЬ: пробираться сквозь чАщу, сквозь зАросли, сквозь тумАн. Всегда несёт на себе побочное удар. и предлог КРОМЕ: собрались все, кроме негО; ничего не надое собой брать, кроме кнИг; кроме берЁз, здесь были и липы. Слабоударяемыми бывают некоторые глагольные формы и вводные слова БЫЛО, БЫВАЛО, СТАЛО БЫТЬ: я хотЕл было взяться за чтение; сидЕли мы, бывало, вечерами вместе и беседовали; Он, стало быть, собрался уехать?
Слабоударяемыми бывают некоторые глагольные формы и вводные слова БЫЛО, БЫВАЛО, СТАЛО БЫТЬ: я хотЕл было взяться за чтение; сидЕли мы, бывало, вечерами вместе и беседовали; Он, стало быть, собрался уехать?
Однако не следует увлекаться побочными ударениями. Если речь диктора выдержана в умеренном темпе, произношение безударных слов — с второстепенным удар., а слабоударяемых слов — с нормальным удар. будет создавать избыточную акцентуацию, которая лишь утяжеляет восприятие речи и мешает слушателям.
Слова с побочным ударением привлекают к себе особое внимание. В большинстве своём каждое самостоятельное слово имеет всего одно удар. Но есть слова с большим количеством слогов и сложные по составу, которые имеют еще и побочное удар. Это главным образом:
слова, образованные путем сложения двух основ: цельнометаллИческий, черепно- мозговОй, бессрочноотпускнОй;
слова с некоторыми префиксами иноязычного происхождения: антиклерикАльный, ультрареакциОнный, супероблОжка, ремилитаризАция;
сложносокращенные слова: земснарЯд, райсовЕт, обллИт, энергосбЫт, партконферЕнция.
Если в слове два ударения, то основное ударение будет располагаться ближе к концу слова, а ближе к началу находится побочное. Также имеет значение и удалённость основного удар. от начала слова: человеконенавИстник, времяпрепровождЕние, хлопкоочистИтельный.
Широко употребляемые сложные слова, если они невелики, обычно не имеют побочного ударения: пылесОс, водостОк, садовОд, водопровОд, чернозЁм, благодАрный, дальнозОркий, землемЕр, многовековОй. Без побочного удар. произносятся и такие распространённые слова, как землетрясЕние, сельскохозЯйственный и другие.
Побочные ударения чаще появляются в книжных по стилистической окраске словах: (клятвопреступлЕние, книгохранИлище) и в специальных словах: (электронновАкуумный, гальваноакУстика, радиорепортАж, киносценАрий, фотокорреспондЕнт, шахтоподъЁмник).
Бывает, при образовании сложного слова путем словосложения основное удар. перемещается ближе к центру слова и оказывается не на том слоге, на который падает в слове, употребленном самостоятельно. Так, мы говорим:
Так, мы говорим:
боЕц — но молотобОец, сваебОец;
волновОй — но коротковолнОвый, длинноволнОвый;
заводскОй — но электрозавОдский;
леснОй — но малолЕсный;
привознОй — но дальнепривОзный;
проводнОй — по электропровОдный;
продавЕц — но книгопродАвец;
сплавнОй — но лесосплАвный;
творЕц — но стихотвОрец;
ушнОй — но околоУшный;
цветнОй — но одноцвЕтный.
Сложные прилагательные и сложные порядковые числительные, в первой части которых есть 3, 4, от 11 до 20, а также 30, 40, 50, 60… 100, нередко произносятся с двумя ударениями (побочное — на числительном): одиннадцатиметрОвый, пятнадцатилитрОвый, семидесятикилометрОвый, девяностолЕтний, стотЫсячный.
С двумя побочными удар. на первой части и с основным ударением на второй части произносятся сложные слова четырёхсотрублЁвый, электромашиностроЕние, автомотогОнщик.
Всегда имеют побочное удар. сложные слова, пишущиеся через дефис: контр-адмирАл, кают-компАния, плащ-палАтка, яхт-клУб.
Побочное ударение может падать на некоторые приставки: СВЕРХ-, ПОСЛЕ-, МЕЖ-. Но и здесь играет роль степень употребительности слова. Например, с побочным удар. произносятся слова послеубОрочный, послеоперациОнный, послеродовОй. Но оно отсутствует у слов послезАвтрашний, послеобЕденный, послеслОвие. А в словах с приставками МЕЖ- к СВЕРХ- побочное удар. ставится всегда: межнационАльный, межотраслевОй, межпланЕтный; сверхглубИнный, сверхмобИльный сверхскоростнОй.
Побочное ударение необходимо, чтобы правильно произносить соответствующие гласные в слове. Если произнести слово хозмАг без второстепенного удар., то оно будет звучать так: хазмАк. Слушатель может не догадаться о значении такого слова. Следовательно, побочное удар. выполняет важную смысловую функцию. Кроме того, оно играет и существенную стилистическую роль. Появление второстепенного удар. там, где оно не обязательно, свидетельствует о просторечном стиле, например: общежИтие, восемьсОт, девятьсОт.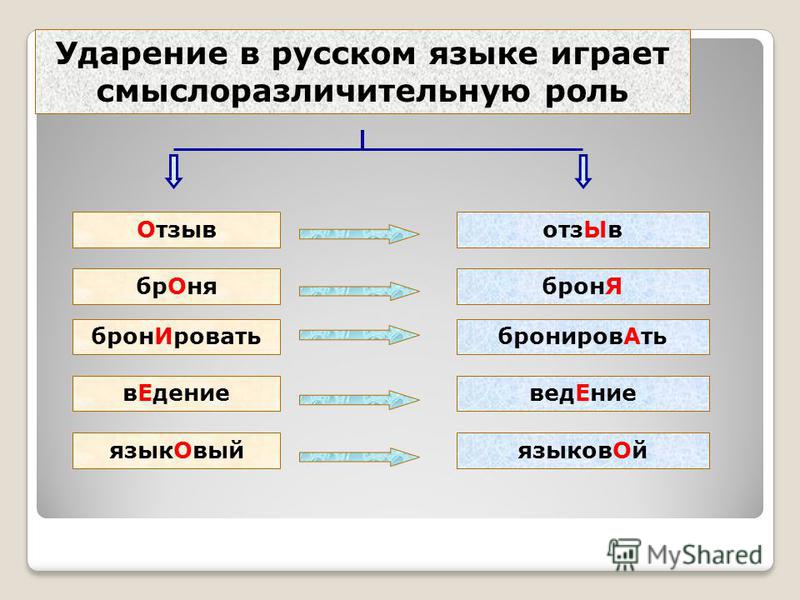 Кроме недопустимой просторечной окраски, такая избыточная ударность придаёт речи утомительность и трудна для восприятия.
Кроме недопустимой просторечной окраски, такая избыточная ударность придаёт речи утомительность и трудна для восприятия.
Изобразительно-выразительные возможности ударения широко используются в публицистических и художественных произведениях. Автор даёт персонажу яркую характеристику, показав своеобразие его удар. и произношения. Проявляются социальная, национальная, профессиональная принадлежность персонажа и степень его образованности. Но, в стилистически нейтральной речи отступление от литературных норм недопустимо. Нормативное словесное ударение способствует правильному восприятию и действенности звучащего слова.
200 слов с правильным ударением, в которых вы, возможно, делаете ошибки
Даже грамотный человек, выучив правила написания слов, может ошибаться в постановке ударений. Правописание некоторые просто зубрят, да и обычно есть время подумать над тем, какую букву поставить в слове. А вот с устной речью ситуация иная – она живая. Вот и появляются самые разные варианты ударений, делая иногда слова и вовсе неузнаваемыми. Давайте разберемся, как правильно произносить некоторые популярные слова, и что стало тому причиной. Заглавной буквой мы выделим ту, на которую и падает ударение в слове.
Вот и появляются самые разные варианты ударений, делая иногда слова и вовсе неузнаваемыми. Давайте разберемся, как правильно произносить некоторые популярные слова, и что стало тому причиной. Заглавной буквой мы выделим ту, на которую и падает ударение в слове.
♥ ПО ТЕМЕ: Как поставить ударение в словах на iPhone, Android, Windows и Mac.
ЖерлО́
Трудно поверить именно в такую постановку ударения. Но так велят словари – ударение надо делать именно на последний слог. А вот во множественном числе ударение сместится на букву «е». Это правило действует как в отношение жерла вулкана, так и в отношении жерла оружия.
ФетИ́ш
Почему ударение тут падает на последний слог – объяснить просто. Слово это пришло из французского языка, а там именно последний слог оказывается под ударением. Наверняка вы слышали где-то об этом, даже изучая английский язык или испанский.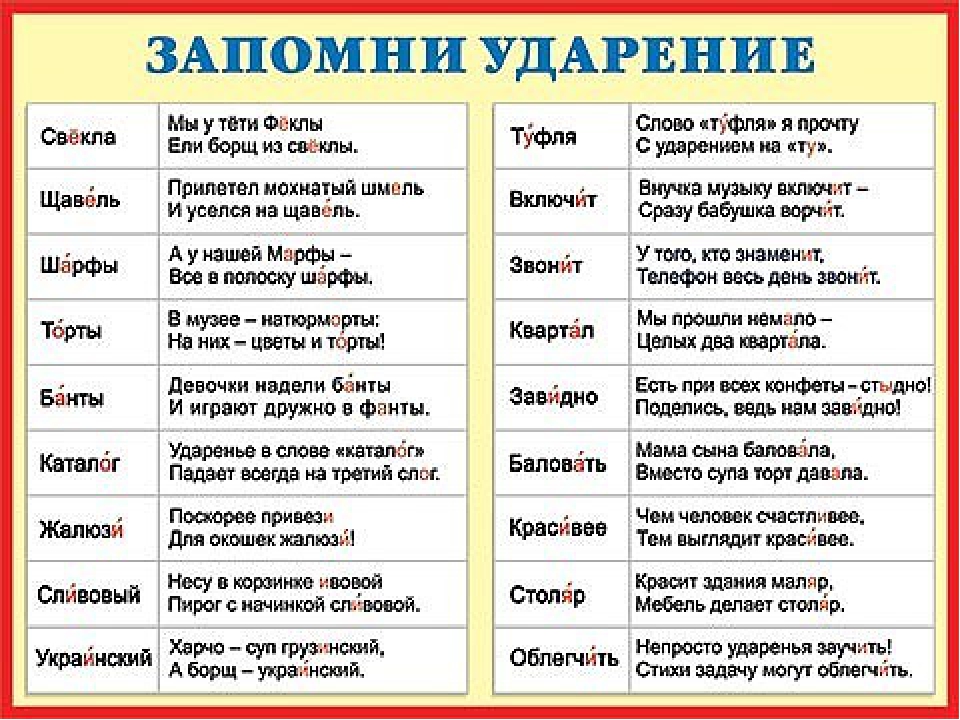 В русском языке оригинальное ударение сохранилось, это даже и хорошо – не придется запоминать очередное исключение из правил. Достаточно лишь помнить происхождение слова.
В русском языке оригинальное ударение сохранилось, это даже и хорошо – не придется запоминать очередное исключение из правил. Достаточно лишь помнить происхождение слова.
ЖалюзИ́
Это слово соответствует упомянутому выше правилу. Жалюзи имеет французское происхождение, стало быть, ударение надо ставить на последний слог.
♥ ПО ТЕМЕ: На этой картинке 16 кругов, вы их видите?
ЧЕ́рпать
Этот глагол происходит от существительного черпак. Такой инструмент предназначен для набора чего-то жидкого или сыпучего. Но с помощью него можно чЕрпать как щебень, так и суп.
СлИ́вовый
Неопытный официант может удивиться тому, что вы заказываете именно слИвовый соус в ресторане. Но только знаток русского языка похвалит вас за верную постановку ударения в этом слове, ведь большинство из нас постоянно делает тут ошибку.
ГрУ́шевый
Давайте закончим с плодами/ягодами. Касающееся производных из сливы правило относится и к грушевым продуктам. Ударение в таких прилагательных тоже надо ставить на первый слог.
Касающееся производных из сливы правило относится и к грушевым продуктам. Ударение в таких прилагательных тоже надо ставить на первый слог.
♥ ПО ТЕМЕ: Говорящий попугай: какие птицы говорят, легче всего учатся и как научить говорить.
СирО́ты
Мало кто знает, что это слово произошло от прилагательного «сирый», которое уже редко и употребляется. Но ударение в слове «сироты» ставить на первый слог просто неграмотно.
НоворождЁнный
Люди часто в этом слове заменяют букве «ё» на «е». Из-за этого легко ошибиться с постановкой ударения. Вот и звучит в повседневной речи слово «новорОжденный». А ведь с «ё» в составе ударный слог очевиден. В русском языке практически всегда интонацией мы делаем акцент на этой букве.
ТанцО́вщица
Предлагаемый словарем вариант выглядит немного скучновато, тогда как в танцовщИце есть нечто задорное, огненное и даже экзотическое. Но все же стоит употреблять слово правильно.
♥ ПО ТЕМЕ: Что можно сделать из старого монитора? 12 идей.
ФлюорогрА́фия
Чего уж стесняться, чаще всего мы вообще заменяем это слово куда более компактным ФЛЮ. Если же кто-то пошлет нас на флюорографИю, то его можно смело поправлять. Обследование лишним не будет, как и знание правильного ударения. А с этим словом даже медики испытывают проблемы.
РефлЕ́ксия
Это слово неправильно произносят даже профессионалы, еще больше увеличивая путаницу. Психологи часто говорят о рефлексИи, но все же стоит поверить в этом вопросе орфографическому словарю.
ПрибылА́, но прИ́были
Этот глагол вполне соответствует правилу постановки ударения на приставку, если слово мужского или среднего рода или на окончание, если оно в женском. А вот во множественном числе ударение приходится на приставку.
♥ ПО ТЕМЕ: Что такое сусеки, почему ножки курьи, а не куриные и из чего сделана лубяная избушка: слова из сказок, о происхождении которых вы могли не знать.
О́трочество
Это слово мы произносим все реже и реже, возвращаясь к нему разве что на уроках литературы. Но все же стоит знать, как правильно называть одну из повестей Льва Толстого.
КровоточА́щий
Если в беседе с настоящим филологом ошибочно поставить ударение в этом слове, то у него явно что-нибудь начнет кровоточить.
КУ́хонный
У каждой семьи есть кухня и что-то кухонное. Но правильно ли мы называем эти вещи? Мойдодыр Чуковского, обидевшись на свое неверное свое произношение, наверняка бы погнался бы за невежами.
♥ ПО ТЕМЕ: 70 интересных фактов об IT-технологиях, о которых вы могли не знать.
ВероисповЕ́дание
Это слово такое длинное и с таким множеством слогов, что ошибиться можно как в его написании, так и в его произношении. Если возникают сомнения, то стоит вспомнить инфинитив глагола «исповЕдовать».
ЗавИ́дно
Чувство зависти возникает у каждого из нас. Неплохо было бы в точности знать, как уметь его описывать.
ОбеспЕ́чение
Существует простое правило, которое позволит легко запомнить правильное ударение в этом слове. Если в нем просматривается печЕнье (обеспечЕнье), то вы сместили акцент явно не в ту сторону.
♥ ПО ТЕМЕ: Как правильно произносить Xiaomi на русском и что обозначает это слово.
ДиспансЕ́р
Никаких диспАнсеров в природе не существует, в этом слове ударение надо ставить на последний слог.
ВручИ́т
Не стоит дожидаться, что вам кто-то врУчит награду за грамотность при использовании такого слова. Так говорить – ошибочно.
ХодА́тайство
Неверный вариант с ходатАйством встречается в нашей речи чуть ли не чаще, чем правильный. Но словарь однозначно указывает, как именно надо употреблять это слово. Ударение следует ставить на второй слог.
♥ ПО ТЕМЕ: Космические скорости — насколько быстро нужно лететь, чтобы покинуть Землю, планетную систему и галактику?
КвартА́л
С этим словом связан один из главных мифов в области ударений в русском языке. Почему-то считается, что при упоминании части года надо говорить квАртал, а при разговоре о части города – квартАл. Но в реальности все проще – ударение следует ставить на последний слог вне контекста использования слова.
ДоговО́р
Это слово очень популярное, что и приводит к постоянным ошибкам в постановке в нем ударений. В бытовой речи еще можно выделять первый слог, ведь такой вариант встречался даже в словарях прошлого века. Но если мы хотим считаться грамотными людьми, то в любых обстоятельствах нужно говорить «договОр».
ВероисповЕ́дание
Статья 28 Конституции Российской Федерации есть статья, которая гарантирует свободу вероисповЕдания. Надо знать не только свои права, но и как правильно произносить их название.
♥ ПО ТЕМЕ: Чем веганы отличаются от вегетарианцев.
ФенО́мен
Здесь мы имеем дело со сложным вопросом. Произношение некоторых слов надо просто выучить наизусть и не задумываться о причинах постановки ударения в это место. Из этого числа как раз «феномен». В словарях можно даже найти разное объяснение употребления слова. «ФенОмен» считается научным термином, а вот при указке на какого-то человека с выдающимися талантами или способностями уже можно говорить как фенОмен, так и феномЕн. Но предпочтительным все же считается вариант с ударением на второй слог в любом значении слова. Вы можете стать настоящим фенОменом, правильно выбирая произношение слов.
КаталО́г
Сегодня каталоги товаров переместились в интернет-магазины, а проблема с произношением осталась прежней. Времена меняются, а слово и в прошлом, и теперь следует произносить правильно – каталОг.
В аэропО́ртах
Любой из путешественников немало времени проводит в аэропортах. Но как правильно произносить это слово? Подход здесь примерно такой же, как и с другими словами во множественном числе: «тортов», «шарфов» и «бантов». Так хочется поставить ударение на последний слог, уж больно естественным это кажется! Но мы хотим вас предостеречь! Правильно говорить: «у аэропОрта», «в аэропОртах». Если же требуется обозначить внутреннее местонахождение, то оно будет в аэропортУ. На конце окажется ударная буква «у».
♥ ПО ТЕМЕ: Лучшие триллеры: Список самых лучших 35 фильмов с описанием, трейлерами и ссылками.
КрасИ́вее
Комплименты любят слушать все, но они должны быть искренними и звучать грамотно. Очевидно, что в прилагательном «красивый» ударение падает на второй слог. Оно сохраняется там и при употреблении в кратких формах и в сравнительной степени – запомнить это просто. А в речи и письме к тому же нельзя ставить «й» в конце этого слова – никакого «красивей».
ОдноврЕ́менно и ОдновремЕ́нно
Наречие одновременно подобно прилагательному одновременный – ударение тут можно ставить и на третий, и на четвертый слог. Оба эти варианта считаются допустимыми и равноправными. Даже драматург Е.Гришковец упомянул об этом явлении в своей пьесе «Одновременно».
Существует словарь «Русское словесное ударение» от М. В. Зарвы, рекомендуемый для сотрудников прессы. Там правильным вариантом называют «одноврЕменно». Аналогичного мнения придерживаются старые словари Даля и Долопчева. А вот в современных «Орфоэпическом словаре русского языка» и «Словаре трудностей русского языка» говорится о двух допустимых формах произношения, разрешив и «одновремЕнно». А вот говорить «одновремЁнно» уж точно не стоить. И если вы принципиально хотите выбрать один вариант, то уж предпочтите канонический «одноврЕменно».
♥ ПО ТЕМЕ: У кого больше всех подписчиков в Инстаграм – 35 самых популярных аккаунтов.
И еще немного распространенных слов:
А
агЕнт
агронОмия
акрОполь
алкогОль
алфавИт
Амфора
анАлог
анАтом
анонИм
апокАлипсис
арАхис
арЕст
аргумЕнт
асимметрИя
астрОлог
астронОм
атмосфЕра
афЕра
аэропОрты, им.п. мн.ч.
Б
бАнты, им.п.мн.ч
бАржА
бОроду, вин.п.ед.ч.
бУнгало
балОванный, прич.
балУясь
баловАть
блАговест
блуднИца
бралА
бралАсь
бухгАлтеров, род.п. мн.н
бюрокрАтия
В
вЕрба
вЕчеря
вОвремя
вОгнутый
валовОй
вандАлы
вдовствО
вернА
вероисповЕдание
вероисповЕдание
ветеринАрия
взАпуски
взапертИ
взялА
взялАсь
включЁн
включЁнный
включИм
включИт
включИшь
влилАсь
водопровОд
воздухопровОд
ворвалАсь
воспринялА
воспроизведЕние
воссоздалА
вручИт
втрИдорога
вчистУю
♥ ПО ТЕМЕ: БелАЗ-75710: 1 300 литров топлива на 100 км и другие 7 фактов о лучшем самосвале мира из Беларуси.
Г
гЕнезис
гЕрбовый
газопровОд
гастронОмия
гегемОния
гипОтеза
гналА
гналАсь
гомеопАтия
граждАнство
граждАнство
грошОвый
Д
дОверху
дОгмат
дОнизу
дОсуха
дОсыта
давнИшний
дефИс
дефИс
диалОг
диспансЕр
диспансЕр
добелА
добралА
добралАсь
довезЁнный
договОр
договорЁнность
дождалАсь
дозИровать
дозвонИтся
дозвонЯтся
докраснА
докумЕнт
докумЕнт
донЕльзя
донЕльзя
досУг
досУг
дотрОнуться
дремОта
духовнИк
Е
евАнгелие
еретИк
еретИк
♥ ПО ТЕМЕ: Что такое карат? Как определить вес драгоценного камня и пробу золота в каратах.
Ж
жалюзИ
жалюзИ, ср.р.и мн.ч.
ждалА
жилОсь
З
зАгнутый
зАгодя
зАгодя
зАнял
зАняло
зАнятый
зАсветло
зАтемно
завИдно
завИдно
завсегдАтай
задОлго
закУпорив
закУпорить
закУпорить
занялА
занятА
запертА
запломбировАть
заселЁн
звалА
звонИм
звонИт
звонИшь
зимОвщик
злОба
знАмение
знАчимость
знАчимый
зубчАтый
И
Издавна
Иконопись
Иксы
Искоса
Искра знания и искрА зажигания
Исстари
Исстари
игУмен
идеОлог
иерОглиф
изОгнутый
избалОванный
избаловАть
издрЕвле
изобретЕние
импЕрский
инАче
инсУльт
инстИнкт
исключИт
искривИться
исчЕрпать
♥ ПО ТЕМЕ: Почему вино сухое, или какие вина называют сухими, а какие сладкими?
К
кАмбала
кАшлянуть
кОнусы,кОнусов
кУхонный
кУхонный
каталОг
каталОг
каучУк
квартАл
квартАл
кедрОвый
киломЕтр
киломЕтр
клАла
клЕить
клЕить
коклЮш
корЫсть
корЫсть
кормЯщий
крАлась
крАны
красИвее
красИвейший
кремЕнь
кремЕнь
кренИтся
кровоточАщий
кровоточИть
Л
лЕкторы,лЕкторов род.п. мн.ч.
лгалА
лилА
лилАсь
ловкА
ломОта
ломОть
лубОчный
лыжнЯ
М
мАркЕтинг (допустимы оба ударения)
мЕльком
мЕстностей род.п. мн.ч
магазИн
мастерскИ
медикамЕнты
метонИмия
мозаИчный
молОх
молЯщий
монолОг
мусоропровОд
мусоропровОд
мытАрство
♥ ПО ТЕМЕ: Почему рождаются рыжие люди и чем они отличаются от остальных.
Н
нАискось
нАчал
нАчали
нАчатые-убрать
нАчатый
нЕдруг
нЕнависть
нЕнецкий
нОвости,новостЕй
нОгтя, род.п ед.ч.
наОтмашь
навЕрх
навралА
наговОр
надОлго
наделИт
надорвалАсь
нажИвший
нажИлся-убрать
нажитА
назвалАсь
назлО
накренИт
налИвший
налилА
налитА
намЕрение
намЕрение
нанЯвшийся
нарОст
нарОст
нарвалА
насорИт
начАв
начАвший
начАвшись
начАть
начАть
началА
недУг
незадОлго
некролОг
некролОг
ненадОлго
ненадОлго
несказАнно
нефтепровОд
низИна
низведЁн
новоприбЫвший
новорождЁнный
О
обеспЕчение
обеспЕчение
обетовАнный
обзвонИт
облегчИт
облегчИть
облегчИть
облилАсь
обнаружЕние
обнялАсь
обогналА
ободрЁн
ободрЁнный
ободрИть
ободрИть
ободрИшься
ободралА
ободренА
обострЁнный
обострИть
объезднОй
одОбренный
одолжИт
озвУчение
озлОбить
озлОбленный
ознакОмленный
оклЕить
окружИт
опОшлят
оперИться
опломбировАть
определЁн
оптОвый
оптОвый
освЕдомить
освЕдомиться, освЕдомишься
осведомлЁнный
остриЁ
осужденА
отбылА
отдАв
отдалА
откУпорил
откУпорить
отключЁнный
отозвалА
отозвалАсь
оторвалА
Отрочество
оценЁнный
♥ ПО ТЕМЕ: Что такое лошадиная сила и сколько лошадиных сил в одной лошади?
П
пАсквиль
пЕтля
пОнял
пОручни
пОстриг
пУстошь
пУстынь
партЕр
патриАрхия
перезвонИт
перекрОенный
перелилА
петлЯ
пиццерИя
плЕсневеть
платО
плодоносИть
пломбировАть
поИмка
повторЁнный
повторИт
поделЁнный
подзаголОвок
поднЯв
подрОстковый
подчистУю
позвалА
позвонИт, позвонИшь
поискОвый
полилА
положИл
положИть
полтергЕйст
понЯв
понЯвший
понялА
портфЕль
послАла
прИбыл
прИбыло
прИкус
прИнял
прИняли
прИнятый
приручЁнный
предвосхИтить
премировАть
прибЫв
прибылА
приговОр
придАное
призЫв
принУдить
принУдить
принЯть
приручЁнный
прогИб
прожИвший
прозорлИва
проторЁнный
процЕнт
процЕнт
псевдонИм
пулОвер
пургА
путепровОд
Р
рАджа
рАпорт
рОвненько
рОзги
развитОй
ракУшка
рвалА
РевЕнь
♥ ПО ТЕМЕ: Откуда взялось выражение (жест) ОК и что в действительности оно означает?
С
сЕтчатый
сОгнутый
сОгнутый
сОздало
сабО
свЁкла
свЁкла
сверлИт
сверлИшь
сегмЕнт
сирОты
слИвовый
снялА
снятА
созЫв
созЫв
создАвший
создАл
создалА
создалА
созданА
сорИт
сосредотОчение
срЕдства,им.п.мн.ч.
срЕдствами
стАтуя
столЯр
столЯр
Т
тАинство
тОртов
тОрты
тОтчас
тУфля
табУ
тамОжня
танцОвщица
твОрОг (двойное ударение)
тигрОвый
толИка
тошнотА
трУбчатый
трубопровОд
У
убралА
убыстрИть
углубИть
углубИть
уговОр
узаконЕние
украИнский
укрепИт
умЕрший
упрОчение
♥ ПО ТЕМЕ: 35 лучших оптических иллюзий, ставших интернет-мемами.
Ф
факсИмиле
фенОмен (необычное явление)
феномЕн (выдающийся человек)
фетИш
флюорогрАфия
Х
ходАтайство
Ц
цЕнтнер
цемЕнт
цепОчка
♥ ПО ТЕМЕ: Скрытый смысл логотипов известных компаний.
Ч
чЕрпать
чИстильщик
Ш
шАрфы
шофЁр
Щ
щЁлкать
щИколотка
щавЕль
щемИт
Э
экспЕрт
Экскурс
электропрОвод
Смотрите также:
Ударение в кратких прилагательных — Русский язык без проблем
вернуться на страницу «Фонетика в таблицах» «Таблицы«, «Фонетический разбор»
Ударение в кратких прилагательных
Наименее устойчиво ударение в кратких формах прилагательных. Это, как правило, слова с односложными основами без суффиксов (или с простейшими древними суффиксами -к-, -н-, частично «поглощенными» основой).
Существует три закономерности в перемещении ударения в этих прилагательных, которые, однако, не всегда выдерживаются, и поэтому в некоторых случаях допустимы колебания.
1) В краткой форме женского рода – ударение на окончании, в остальных формах – на основе. Допустимы колебания в формах среднего рода и множественного числа (типы 1-3).
2) Во всех формах – ударение на основе. Допустимы колебания в форме женского рода (тип 4).
3) В краткой форме мужского рода ударение на основе, в остальных формах – на окончании (тип 5).
формы с суффиксами –ив, –лив, –чив, –им, –н, –альн, –ельн, –ист. ударение падает на тот же слог, что и в прилагательных полной формы
красИвый – красИв, красИва, говорлИвый – говорлИв, говорлИва, устОйчивый – устОйчив, устОйчива, немЫслимый – немЫслим, немЫслима, грАмотный – грАмотен, грАмотна,
слова с односложными основами без суффиксов, или с суфф. -к, -н. в краткой форме прилагательного женского рода ударение перемещается на окончание
бЫстрый – бЫстр, быстрА, гОдный – гОден, годнА, густОй – гУст, густА, гИбкий – гИбок, гибкА, грЕшный – грЕшен, грешнА
Если в краткой форме женского рода ударение падает на окончание, то в сравнительной форме оно оказывается на суффиксе –ее: длиннА – длиннЕе, виднА – виднЕе.
Если же в краткой форме женского рода ударение стоит на основе, то и в сравнительной степени оно остаётся на основе: красИва — красИвее, ленИва — ленИвее
Колебания ударения допускаются в следующих кратких прилагательных, относящихся к среднему роду бЕлы – белЫ, велИки – великИ, вИдны – виднЫ, врЕдны – вреднЫ, высОко – высокО, глубОко – глубокО, гОдны – годнЫ, мИлы – милЫ, мАлы – малЫ, нОвы – новЫ, нУжны – нужнЫ, пОлны – полнЫ, стАры – старЫ, ширОки – широкИ.
Перечень прилагательных в краткой форме множественного числа, имеющих нормативное ударение (на основе) блЕдны, блИзки, бОйки, бУрны, вЕрны, врЕдны, глУпы, гОрьки, грЯзны, длИнны, жАлки, знОйны, крАсны, крУглы, лОжны, нИзки, прАвы, прОсты, пУсты, рЕдки, рЕзки, сУхи, Узки, чИсты, Ярки, Ясны.
Остались вопросы — задай в обсуждениях https://vk.com/board41801109
Усвоил тему — поделись с друзьями.
Тесты на тему Ударение
вернуться на страницу «Фонетика в таблицах» «Таблицы«, «Фонетический разбор«
Чередование О-А в корне слова — Уроки Русского
В ряде корней их правописание зависит от ударения.
| Правописание зависит от ударения | ||
| Корень | Правило | Исключения |
| -клан-/-клон- | Без ударения – О; Под ударением – та гласная, что слышится | |
| -гар-/-гор- | изгарь, пригарь, выгарки | |
| -твар-/-твор-
| утварь | |
| -зар-/-зор- | Без ударения — А | зоревать |
| -плав-/-плов- | пловец, пловчиха | |
В корнях –клан-/-клон-, -гар-/-гор-, -твар-/-твор— без ударения пишется буква О, под ударением та гласная, что слышится. Например:
накл..ниться – наклониться; кланяться
Наклониться, поставим ударение: наклонИться. Ударение падает не на корень. Значит, без ударения пишется корень –клон-.
Кланяться. Корень стоит под ударением, пишем букву А.
Загоратьть, поставим ударение: загорАть. Ударение падает не на корень. Значит, без ударения пишется корень –гор-.
заг..рать – загорать; загар
Загар. Корень стоит под ударением, пишем букву А.
творение, поставим ударение: творЕние. Ударение падает не на корень. Значит, без ударения пишется корень –твор-.
тв..рение – творение; творчество
Творчество. Корень стоит под ударением, пишем букву О.
Исключения из этого правила — слова: изгарь, пригарь, выгарки; утварь.
В корнях –зар/-зор-, -плав-/плов— без ударения пишется буква А, под ударением та гласная, что слышится. Например:
З..ря – заря; зорька
Заря, поставим ударение: зарЯ. Ударение падает не на корень. Значит, без ударения пишется корень –зар-.
Зорька. Корень стоит под ударением, пишем букву О.
Попл..вок – поплавок; плавание
Поплавок, поставим ударение: поплавОк. Ударение падает не на корень. Значит, без ударения пишется корень –плав-.
Плавание. Корень стоит под ударением, пишем букву А.
Исключение из этого правила — слова: зоревать; пловец, пловчиха.
Правописание зависит от конечной согласной корня:
| Правописание зависит от конечной согласной корня | ||
| Корень | Правило | Исключения |
| -лож-/-лаг- | ||
| -раст-/-ращ-/-рос- | отрасль, росток, ростовщик, Ростов, Ростислав, на вырост, выросток | |
| -скак-/-скоч-
| скачок, скачу | |
В корнях -лож-/-лаг-, -раст-/-ращ-/-рос-, -скак-/-скоч- правописание гласных зависит от конечной согласной корня. Эти корни надо запомнить. Например:
сл..гаемое – слагаемое
Слагаемое. Корень заканчивается на «г», значит, пишется корень –лаг-.
р..стение – растение
Растение. Корень заканчивается на «ст», значит, пишется корень –раст-.
Подскочить. Корень заканчивается на «ч», значит, пишется корень –скоч-.
подск..чить – подскочить
Исключения из этого правила — слова: отрасль, росток, ростовщик, Ростов, Ростислав, на вырост, выросток; скачок, скачу.
Правописание зависит от суффикса –а-:
| Правописание зависит от суффикса –а- | |
| Корень | Правило |
| -кос-/-кас- | -кас- + -а- -кос- |
В корне –кос-/-кас- пишется А, если после корня следует суффикс –а-. Пишется О, если после корня нет суффикса –а-. Например:
к..сание – касание
Касание. После корня есть суффикса –а-. Значит пишется корень –кас—
Правописание зависит от значения:
| Правописание зависит от ударения | ||
| Корень | Правило | Исключения |
| -мок-/-мак- | -мак-: ‘погружать, опускать во что-то’ -мок-: ‘пропускать, впитывать жидкость, становиться мокрым, влажным’ | |
| -ровн-/-равн- | -равн-: ‘одинаковый, равный, такой же, наравне’ -ровн-: ‘прямой, без изгибов; гладкий, без возвышенностей’ | равнина, поровну |
В корнях -мок-/-мак-, -ровн-/-равн- правописание гласных зависит от значения.
Пишется -мак— в значении ‘погружать, опускать во что-то’. Пишется -мок- в значении ‘пропускать, впитывать жидкость, становиться мокрым, влажным’. Например:
обм..кнуть блин в варенье – обмакнуть.
Обмакнуть блин в варенье. Значение ‘погружать, опускать во что-то’. Значит, пишется корень –мак-.
нам..кать под дождем — намокать
Намокать под дождём. Значение ‘становиться мокрым, влажным’. Значит, пишется корень –мок.
Пишется -равн— в значении ‘одинаковый, равный, такой же, наравне’. Пишется -ровн- в значении ‘прямой, без изгибов; гладкий, без возвышенностей’. Например:
ср..внять условия — сравнять
Сравнять условия. Значение ‘одинаковый, равный, такой же’. Значит, пишется корень –равн-.
пор..внять грядку — поровнять
Поровнять грядку. Значение ‘гладкий, без возвышенностей’. Пишется корень –ровн-.
Исключения из этого правила — слова: равнина, поровну.
Повторим
| Правописание зависит от ударения | ||
| Корень | Правило | Исключения |
| -клан-/-клон- | Без ударения – О; Под ударением – та гласная, что слышится | |
| -гар-/-гор- | изгарь, пригарь, выгарки | |
| -твар-/-твор-
| утварь | |
| -зар-/-зор- | Без ударения — А | зоревать |
| -плав-/-плов- | пловец, пловчиха | |
| Правописание зависит от конечной согласной корня | ||
| -лож-/-лаг- | ||
| -раст-/-ращ-/-рос- | отрасль, росток, ростовщик, Ростов, Ростислав, на вырост, выросток | |
| -скак-/-скоч-
| скачок, скачу | |
| Правописание зависит от суффикса –а- | ||
| -кос-/-кас- | -кас- + -а- -кос- | |
| Правописание зависит от ударения | ||
| -мок-/-мак- | -мак-: ‘погружать, опускать во что-то’ -мок-: ‘пропускать, впитывать жидкость, становиться мокрым, влажным’ | |
| -ровн-/-равн- | -равн-: ‘одинаковый, равный, такой же, наравне’ -ровн-: ‘прямой, без изгибов; гладкий, без возвышенностей’ | равнина, поровну |
В корнях –клан-/-клон-, -гар-/-гор-, -твар-/-твор— без ударения пишется буква О, под ударением та гласная, что слышится.
Исключения: изгарь, пригарь, выгарки; утварь.
В корнях –зар/-зор-, -плав-/плов— без ударения пишется буква А, под ударением та гласная, что слышится.
Исключениея: зоревать; пловец, пловчиха.
В корнях -лож-/-лаг-, -раст-/-ращ-/-рос-, -скак-/-скоч- правописание гласных зависит от конечной согласной корня.
Исключения: отрасль, росток, ростовщик, Ростов, Ростислав, на вырост, выросток; скачок, скачу.
В корне –кос-/-кас- пишется А, если после корня следует суффикс –а-. Пишется О, если после корня нет суффикса –а-.
В корнях -мок-/-мак-, -ровн-/-равн- правописание гласных зависит от значения.
Пишется -мак— в значении ‘погружать, опускать во что-то’. Пишется -мок- в значении ‘пропускать, впитывать жидкость, становиться мокрым, влажным’.
Пишется -равн— в значении ‘одинаковый, равный, такой же, наравне’. Пишется -ровн- в значении ‘прямой, без изгибов; гладкий, без возвышенностей’.
Исключения: равнина, поровну.
Напряжение, деформация и модуль Юнга
Напряжение
Напряжение — это отношение приложенной силы F к площади поперечного сечения –, определяемой как « силы на единицу площади ».
- растягивающее напряжение — напряжение, которое имеет тенденцию к растяжению или удлинению материала — действует нормально по отношению к напряженной области
- сжимающее напряжение — напряжение, которое имеет тенденцию к сжатию или укорачиванию материала — действует нормально по отношению к напряженной области
- напряжение сдвига — напряжение, которое имеет тенденцию к сдвигу материала — действует в плоскости к напряженной области под прямым углом к напряжению сжатия или растяжения
Напряжение растяжения или сжатия — нормальное напряжение
Напряжение растяжения или сжатия, нормальное к плоскости обычно обозначается как « нормальное напряжение » или « прямое напряжение » и может быть выражено как
σ = F n / A (1)
где
σ = нормальное напряжение (Па (Н / м 2 ), фунт / кв. дюйм (фунт f / дюйм 2 ))
F n = нормальная сила, действующая перпендикулярно площади (Н, фунт f )
A = площадь (м 2 , дюйм 2 )
- кип — британская система мер единица силы — равна 1000 фунтов f (фунт-сила)
- 1 кип = 4448.2216 Ньютонов (Н) = 4.4482216 килограммов Ньютонов (кН)
Нормальная сила действует перпендикулярно площади и возникает всякий раз, когда внешние нагрузки имеют тенденцию толкать или тянуть два сегмента тела.
Пример — Растягивающая сила, действующая на стержень
Сила 10 кН действует на круглый стержень диаметром 10 мм . Напряжение в стержне можно рассчитать как
σ = (10 10 3 Н) / (π ((10 10 -3 м) / 2) 2 )
= 127388535 (Н / м 2 )
= 127 (МПа)
Пример — Сила, действующая на квадратную стойку из пихты Дугласа
Сжимающая нагрузка 30000 фунтов действует на короткий квадрат 6 x 6 дюймов столб из пихты Дугласа.Размер стойки в оправе составляет 5,5 x 5,5 дюйма , а напряжение сжатия можно рассчитать как
σ = (30000 фунтов) / ((5,5 дюйма) (5,5 дюйма) )
= 991 (фунт / дюйм 2 , psi)
Напряжение сдвига
Напряжение, параллельное плоскости, обычно обозначается как «напряжение сдвига » и может быть выражено как
τ = F p / A (2)
где
τ = напряжение сдвига (Па (Н / м 2 ), фунт / кв. Дюйм (фунт f / дюйм 2 ))
F p = поперечная сила в плоскости области (Н, фунт f )
A = площадь (м 2 , в 2 )
Поперечная сила лежит в плоскости области и возникает, когда внешние нагрузки имеют тенденцию вызывать два сегмента тела скользить друг по другу.
Деформация (деформация)
Деформация определяется как «деформация твердого тела под действием напряжения».
- Нормальная деформация — удлинение или сжатие отрезка линии
- Деформация сдвига — изменение угла между двумя отрезками прямой, первоначально перпендикулярными
Нормальная деформация и может быть выражена как
ε = dl / l o
= σ / E (3)
, где
dl = изменение длины (м, дюйм)
l o = начальная длина (м, дюйм)
ε = деформация — без единиц
E = Модуль Юнга (модуль упругости) (Па, (Н / м 2 ), фунт / кв. дюйм (фунт f / дюйм 2 ))
- Модуль Юнга
- можно использовать для прогнозирования удлинения или сжатия объекта при воздействии силы.
Обратите внимание, что деформация является безразмерной единицей, поскольку это отношение двух длин.Но также общепринято указывать это как отношение двух единиц длины — например, м / м или дюйм / дюйм .
Пример — напряжение и изменение длины
Стержень в приведенном выше примере имеет длину 2 м и изготовлен из стали с модулем упругости 200 ГПа (200 10 9 Н / м 2 ) . Изменение длины можно рассчитать, преобразовав (3) в
dl = σ l o / E
= (127 10 6 Па) (2 м) / (200 10 9 Па)
= 0.00127 м
= 1,27 мм
Энергия деформации
Напряжение объекта сохраняет в нем энергию. Для осевой нагрузки запасенная энергия может быть выражена как
U = 1/2 F n dl
, где
U = энергия деформации (Дж (Н · м), фут-фунт)
Модуль Юнга — модуль упругости (или модуль упругости) — закон Гука
Большинство металлов деформируются пропорционально приложенной нагрузке в диапазоне нагрузок.Напряжение пропорционально нагрузке, а деформация пропорциональна деформации в соответствии с законом Гука .
E = напряжение / деформация
= σ / ε
= (F n / A) / (дл / л o ) ( 4)
, где
E = модуль Юнга (Н / м 2 ) (фунт / дюйм 2 , psi)
Модуль упругости или модуль Юнга обычно используется для металлов и металлических сплавов и выражается в единицах 10 6 фунтов f / дюйм 2 , Н / м 2 или Па .Модуль упругости при растяжении часто используется для пластмасс и выражается в единицах 10 5 фунтов f / дюйм 2 или ГПа.
Модуль упругости при сдвиге — или модуль жесткости
G = напряжение / деформация
= τ / γ
= (F p / A) / (с / d) (5)
, где
G = модуль упругости при сдвиге — или модуль жесткости (Н / м 2 ) (фунт / дюйм 2 , psi)
τ = напряжение сдвига ((Па) Н / м 2 , psi)
γ = мера деформации сдвига без единицы измерения
p = сила, параллельная граням, на которые они действуют
A = площадь (м 2 , в 2 )
s = смещение граней (м, дюйм)
d = ди положение между смещенными гранями (м, дюйм)
Объемный модуль упругости
Объемный модуль упругости — или объемный модуль — является мерой сопротивления вещества равномерному сжатию.Объемный модуль упругости — это отношение напряжения к изменению объема материала, подвергающегося осевой нагрузке.
Модули упругости
Модули упругости для некоторых распространенных материалов:
| Материал | Модуль упругости — E — | Модуль упругости — G — | Модуль упругости — | ||
|---|---|---|---|---|---|
| (ГПа) (10 6 фунтов на кв. Дюйм) | (ГПа) (10 6 фунтов на кв. ) | ||||
| Алюминий | 70 | 24 | 70 | ||
| Латунь | 91 | 36 | 61 | ||
| Медь | 55 | 23 | 37 | ||
| Железо | 91 | 70 | 100 | Свинец | 16 | 5.6 | 7,7 |
| Сталь | 200 | 84 | 160 |
Механика материалов: напряжение »Механика тонких конструкций
Добро пожаловать в Механику материалов. Этот курс основан непосредственно на основах, которые мы изучили в статике, — вычислении статического равновесия различных конструкций при различных нагрузках. В статике мы рассматриваем внешних сил , действующих на твердых тел .В действительности все тела деформируемы, и эти внешние силы создают внутренних напряжения . Ну тогда что за стресс?
Напряжение — это мера внешней силы , действующей на площади поперечного сечения объекта. Напряжение имеет единицы силы на площадь: Н / м 2 (СИ) или фунт / дюйм 2 (США). Единицы СИ обычно называют паскалями, сокращенно Па . Поскольку 1 Па является неудобно малым по сравнению с напряжениями, которые испытывает большинство конструкций, мы часто будем сталкиваться с 10 3 Па = 1 кПа (килопаскаль), 10 6 Па = МПа (мегапаскаль) или 10 9 Па = ГПа (гигапаскаль).
Существует два типа напряжения, которое может испытывать конструкция: 1. Нормальное напряжение и 2. Напряжение сдвига . Когда сила действует перпендикулярно (или «нормально») к поверхности объекта, она вызывает нормальное напряжение. Когда сила действует параллельно поверхности объекта, возникает напряжение сдвига.
Рассмотрим светильник, подвешенный к потолку на веревке. Поперечное сечение веревки круглое, а вес света тянется вниз перпендикулярно веревке.Эта сила создает в канате нормальное напряжение .
Хорошо, как мы пришли к этому уравнению. За кадром существует множество предположений. На протяжении всего этого курса мы будем предполагать, что все материалы однородны, изотропны и эластичны. Мы также предположим, что объект является «призматическим» — это означает, что поперечные сечения одинаковы по всей его длине (например, огурец является призматическим, а тыквенный орех — нет). Все эти предположения позволяют утверждать, что объект будет деформировать равномерно в каждой точке своего поперечного сечения.Нормальное напряжение в точке поперечного сечения определяется как (с аналогичными уравнениями в направлениях x и y ). :
На каждый небольшой участок поперечного сечения действует одинаковая сила, и сумма всех этих сил должна равняться внутренней равнодействующей силе P . Если мы позволим ΔA перейти к dA, а ΔF перейти к dF, то мы сможем просто проинтегрировать обе части уравнения, и мы придем к нашему соотношению для нормального напряжения.
Это соотношение для нормального напряжения более точно соответствует среднему нормальному напряжению , поскольку мы усреднили внутренние силы по всему поперечному сечению.
Понятие стресса часто бывает трудно понять, потому что его нелегко заметить. Оказывается, размещение прозрачного объекта через кросс-поляризованный свет позволяет непосредственно наблюдать напряжение внутри материала на основе концепции, называемой фотоупругостью:
Напряжение действительно может существовать в материале при отсутствии приложенной нагрузки. Это называется остаточным напряжением, и его можно использовать как способ упрочнения материалов, например, при изготовлении японского меча катана.И наоборот, нежелательные остаточные напряжения могут стимулировать рост трещин и привести к разрушению, как, например, при обрушении Серебряного моста в Западной Вирджинии в 1967 году. Возможно, самый яркий пример остаточного напряжения связан с быстрым охлаждением расплавленного стекла, известным как « Капля принца Руперта »:
Давайте посмотрим на другой пример. Рассмотрим болт, соединяющий две прямоугольные пластины, и растягивающее усилие, перпендикулярное болту. Из диаграммы свободного тела мы видим, что приложенная извне сила оказывает силу, параллельную круглому поперечному сечению болта.Эта внешняя сила приводит к напряжению сдвига внутри болта.
Теперь формальные определения напряжения сдвига принимают форму, аналогичную описанным выше. Рассмотрим напряжение сдвига, действующее на поверхность z элемента:
Напряжение сдвига — это касательное напряжение, действующее по касательной к поперечному сечению, и оно принимает среднее значение:
Важно отметить, что напряжения, которые мы только что описали, составляют средних напряжения .Мы предположили, что вся внешняя сила была равномерно распределена по площади поперечного сечения конструкции — это не всегда так, и мы будем пересматривать это предположение на протяжении всего курса.
Когда вы смотрите на элемент при сдвиге, все кажется немного сложнее. Рассмотрим небольшой кубический элемент внутри конструкции при сдвиге, как показано ниже.
Теперь равновесие требует, чтобы напряжение сдвига, действующее на τ zy , сопровождалось напряжениями сдвига в других плоскостях.Но давайте рассмотрим равновесие сил в направлении y . Зная, что силу можно записать как напряжение (тау), умноженное на площадь (ΔxΔy), мы можем записать это силовое равновесие как:
Поскольку площади куба по определению одинаковы, это означает, что τ zy = τ ‘ zy . Аналогичное силовое равновесие в направлении z -направление приводит к τ yz = τ ‘ yz . Рассмотрим моментное равновесие относительно оси x . Зная, что мы можем записать силу, как и раньше, а плечо момента будет Δz, этот баланс моментов станет:
Это простое соотношение говорит нам, что τ zy = τ yz, и, следовательно, все четыре касательных напряжения имеют равные величины и должны указывать навстречу или от друг друга на противоположных краях элемента.Это соотношение известно как «чистый сдвиг».
1,2 Фактор безопасности
Инженеры используют стресс при проектировании конструкций. Внешняя нагрузка и геометрия конструкции говорят нам, какое напряжение действует в материале, но ничего не говорят нам о самом материале. У каждого материала есть предельное напряжение — мера того, какое напряжение может выдержать материал перед разрушением. Чтобы правильно спроектировать безопасную конструкцию, нам необходимо убедиться, что приложенное напряжение от внешней нагрузки никогда не превышает предельное напряжение материала. Отчасти сложность этой задачи заключается в том, что мы не всегда точно знаем, какова внешняя нагрузка — она может изменяться непредсказуемо, и конструкции, возможно, придется выдерживать неожиданно высокие нагрузки. Чтобы учесть эту неопределенность, мы включили в нашу конструкцию коэффициент безопасности . Коэффициент безопасности — это просто отношение разрушающей нагрузки или напряжения к допустимой нагрузке или напряжению. Разрушение или предельное значение — это свойство материала , в то время как допустимое значение определяется внешней силой и геометрией конструкции .
Сводка
В этой лекции мы представили понятие стресса. Напряжение — это мера того, что материал ощущает от приложенных извне сил. Это просто отношение внешних сил к площади поперечного сечения материала. Силы, приложенные перпендикулярно поперечному сечению, составляют нормальных напряжений , в то время как силы, приложенные параллельно поперечному сечению, составляют напряжения сдвига .Хотя представленные здесь концепции не слишком чужды, большая часть трудностей с этим материалом связана с проблемой правильного расчета статического равновесия . Расчет статического равновесия скажет нам величину и направление приложенных сил, которые мы затем можем использовать для расчета напряжений. Если следующие примеры видео и домашнее задание вызывают у вас затруднения, сейчас самое время вернуться и просмотреть некоторые концепции из вашего курса статики.
Этот материал основан на работе, поддержанной Национальным научным фондом в рамках гранта No.1454153. Любые мнения, выводы, выводы или рекомендации, выраженные в этом материале, принадлежат автору (авторам) и не обязательно отражают точку зрения Национального научного фонда.
Сравнение транскриптомов стрессовых гранул и Р-тел показывает, что трансляция играет основную роль в разделении РНК
ВВЕДЕНИЕ
Эукариотические клетки содержат ряд РНК-белковых сборок, вместе называемых гранулами РНП.Гранулы РНП встречаются в ядре, включая тельца Кахаля, параспеклы и ядрышко, а также в цитоплазме, включая стрессовые гранулы (SG) и Р-тельца (PB). PB представляют собой составные сборки нетранслирующих комплексов мРНК с компонентами аппарата распада РНК (1), которые могут увеличиваться во время стресса (2, 3). SGs также состоят из нетранслирующих РНК, но образуются или становятся легко видимыми только во время стрессовой реакции, когда большое количество мРНК прекращает трансляцию (4-6).
Хотя давно известно, что SG и PB содержат РНК и белок, точный состав РНК и белков этих ансамблей остается загадкой из-за отсутствия подходящих методов очистки.Недавно были использованы три различные методологии для выяснения транскриптомов стрессовых гранул или Р-телец для определения факторов, которые влияют на разделение мРНК на эти гранулы РНП. В частности, Р-тельца были очищены и секвенированы с помощью дифференциального центрифугирования и сортировки активированных флуоресценцией частиц из нестрессированных клеток HEK293, выявив составляющие компоненты PB (7). Напротив, стрессовые гранулы были очищены от подвергнутых стрессу клеток U-2 OS и секвенированы с помощью дифференциального центрифугирования с последующей иммуноочисткой (8, 9).Наконец, в другом исследовании изучали состав гранул РНК только с помощью дифференциального центрифугирования (10). В этом исследовании РНК, осаждающиеся во время дифференциального центрифугирования, назывались осадком РНК-гранул (RG), что показало некоторое сходство с иммуноочищенным транскриптомом SG (8, 10).
Здесь мы обращаемся к трем вопросам очистки и анализа транскриптомов SG и PB. Во-первых, используя один тип клеток и последовательный вычислительный конвейер, мы демонстрируем, что транскриптом RG в стрессовых условиях похож на транскриптом SG, но содержит дополнительные загрязняющие РНК.Во-вторых, наш анализ утверждает, что транскриптом PBs резко сдвигается при стрессе, предполагая, что нацеливание мРНК на PB в первую очередь определяется их статусом трансляции. Наконец, мы демонстрируем, что состав РНК PBs и SGs подобны во время стресса. Эти наблюдения предполагают, что мРНК из одних и тех же генов могут разделяться как на SG, так и на PB, и что различия в составе мРНК и / или связанных с ними белков диктуют специфичность ассоциации гранул RNP.
РЕЗУЛЬТАТЫ
Характеристика транскриптома осадка гранул нестрессированной РНК.Транскриптом гранул РНК (RG) недавно был охарактеризован с помощью дифференциального центрифугирования и секвенирования транскриптома (RNA-Seq), и он был определен как популяция РНК, осаждающихся при стрессе и без него (10). Если простое гранулирование сборок RNP может дать точное представление о транскриптоме Р-тельца или стрессовых гранул, это будет дешевле, быстрее и потребует меньшего количества клеток, чем более сложные методы иммуноочистки (7, 8). Хотя анализ транскриптома РНК-гранул исключительно с помощью дифференциального центрифугирования был проведен в клетках фибробластов мыши, мы хотели изучить, как аналогичная методология выполняется в линии клеток человека (U-2 OS) и насколько хорошо осажденная фракция РНП коррелирует с транскриптомом SG. определяется путем иммуноочистки из той же клеточной линии (8).Таким образом, аналогично предыдущим методам (10), мы выделили фракцию тяжелой РНП с помощью дифференциального центрифугирования и проанализировали РНК в этих фракциях с помощью и без стресса с помощью RNA-Seq.
Сначала мы проанализировали таблетку RG в отсутствие напряжения. В этом препарате мы сначала удалили ядра центрифугированием (1000 × г ). После этого начального вращения мы выделили РНК, которые осаждались при 16000 × г , и сравнили эту популяцию РНК с полной РНК с истощенным ядром. Библиотеки последовательностей РНК из нестрессированных клеток воспроизводились как для осадка RG, так и для фракций тотальной РНК, обедненных ядром (рис.1А и Б). Тройные повторы тотальной РНК имели тенденцию иметь большее сходство друг с другом, чем тройные повторы РНК осадка RG, что позволяет предположить, что осадок RG содержит другой поднабор РНК, чем тотальная РНК (рис. 1С). Однако мы отмечаем, что различия между тотальной РНК и осадком RG были небольшими, предполагая, что нестрессированный осадок RG имеет транскриптом, сходный с транскриптомом цитозольного транскриптома. В соответствии с аналогичной методологией, показатели обогащения осадка RG без стресса положительно коррелировали с ранее выделенным осадком RG без стресса из фибробластов мыши ( R = 0.345) (рис. 1D).
Рис. 1.
Транскриптомы осадка гранул без стресса РНК воспроизводимы. (A) Трехмерная (3D) диаграмма рассеяния, изображающая нормализованное количество считываний из библиотек РНК-seq из трех экземпляров осадка гранул нестрессированной РНК. (B) Трехмерная диаграмма рассеяния, изображающая нормализованное количество считываний из библиотек RNA-seq из трех повторений нестрессированной общей РНК. (C) Таблица, изображающая парные коэффициенты корреляции Пирсона между осадками гранул РНК и общей РНК в нестрессированных клетках. (D) Диаграмма рассеяния нестрессированного обогащения RG, полученного из фибробластов мыши, в сравнении с обогащением нестрессированным RG в клетках U-2 OS.
Поскольку существуют различия между РНК осадка и общей цитозольной РНК, мы стремились определить, какие транскрипты были по-разному обогащены внутри осадка и истощены из осадка. Анализ дифференциального обогащения выявил многочисленные транскрипты, которые были значительно ( P <0,01) обогащены ( n = 2161) или истощены ( n = 1304) осадка (рис. 2A). Онтология генов для обогащенных транскриптов выявила множество транскриптов, участвующих в метаболических процессах (рис.2Б). Онтология генов истощенных транскриптов идентифицировала многие транскрипты, кодирующие белки, которые участвуют в нацеливании на мембрану эндоплазматического ретикулума (ER) (Fig. 2B). Это согласуется с ER, остающимся в супернатанте при 16000 × г (11). Митохондрии должны оседать при вращении 16000 × г (11). Действительно, мы наблюдаем, что транскрипты, кодируемые митохондриями, представляют собой некоторые из наиболее высоко экспрессируемых транскриптов, которые обогащены этой методологией (Fig. 2C). Таким образом, нестрессированный транскриптом осадка РНК лишен РНК, связанной с мембранами, и обогащен РНК, локализованными в митохондриях или кодирующими метаболические ферменты.
FIG 2
Характеристика осадка гранул нестрессированной РНК. (A) График МА, отображающий значения log 2 кратного изменения (ненапряженный осадок RG / ненапряженная общая РНК) в зависимости от численности (количество фрагментов на килобаз на миллион [FPKM]). Гены обозначены цветом в зависимости от их значения. Значимые гены ( P <0,01) окрашены в красный цвет, а незначимые ( P > 0,01) — в синий. (B) Анализ онтологии генов для обогащенных и истощенных транскриптов. (C) Увеличенное изображение диаграммы рассеяния, подчеркивающей положение митохондриальных транскриптов.(D) График в виде прямоугольников, изображающий длину транскрипта для обогащенных RG и истощенных RG транскриптов как в стрессированных, так и в нестрессированных клетках. (E) График в виде прямоугольников, изображающий значения эффективности трансляции (18) для RG-обогащенных и RG-истощенных транскриптов в нестрессированных клетках.
Мы стремились изучить показатели, которые могут играть роль в определении того, обогащена ли РНК по-разному в ненапряженном осадке. Мы и другие (7, 8, 10, 12) ранее определили длину трансляции и транскрипта как два преобладающих показателя, которые коррелируют с локализацией РНК в цитоплазматических ансамблях, таких как Р-тельца и стрессовые гранулы.Сначала мы проверили, коррелирует ли длина транскрипта с обогащением осадка. В соответствии с наблюдениями в стрессовых гранулах и P-теле, длинные РНК также имеют тенденцию накапливаться в гранулах в отсутствие стресса (Fig. 2D). Однако смещение длины гораздо менее выражено, чем смещение длины, наблюдаемое в гранулах напряжения (8). Таким образом, длина играет некоторую роль в определении состава РНК фракции осадка RG даже в условиях отсутствия стресса.
Затем мы проверили, существует ли смещение трансляции между транскриптами РНК, обогащенными осадком, и транскриптами РНК, обедненными осадком.Мы не увидели значительного смещения эффективности трансляции при сравнении обогащенных гранулами и обедненных гранулами транскриптов (рис. 2E). Это контрастирует со стрессовыми гранулами и Р-тельцами, которые оба склонны нести плохо транслируемые транскрипты (7, 8, 12). Это различие, однако, согласуется с генной онтологической идентификацией метаболических генов в осадке гранул РНК, которые обычно являются хорошо транслируемыми генами (рис. 2B).
Взятые вместе, наши результаты показывают, что субпопуляция RNPs осаждается в нестрессовых условиях.Транскрипты, которые осаждаются, имеют тенденцию быть длинными и / или имеют тенденцию кодировать гены, участвующие в метаболизме, или гены, кодирующие белки, нацеленные на митохондрии, в то время как транскрипты, которые не осаждаются, имеют тенденцию быть короче и / или кодируют гены, которые локализуются в митохондриях. ER мембрана.
Характеристика транскриптома осадка стрессовых гранул РНК. Ранее было показано, что осадок гранул стрессовых РНК имеет состав РНК, подобный составу стрессовых гранул, выделенных с помощью иммуноочистки (10).В этом предыдущем исследовании авторы отметили, что некоторые из РНК, обогащенных осадком RG, подвергнутого стрессу, были теми же РНК, которые осаждались в условиях отсутствия стресса. Таким образом, мы хотели изучить, как осадок подвергнутой стрессу RG соотносится с осадком РНК, подвергнутым стрессу. Чтобы выделить подвергнутый стрессу осадок RG, мы обрабатывали клетки U-2 OS 0,5 мМ арсенита натрия в течение 1 ч, а затем таким же образом секвенировали обедненную ядром общую цитоплазматическую РНК и гранулированные РНП. Библиотеки
RNA-seq были воспроизводимы как для троекратных повторов тотальной РНК с вычитанием ядра, так и для троекратных повторов осадка RG (рис.3А и В). РНК, которые осаждались в стрессовых условиях, существенно отличались от стрессированной общей цитозольной РНК (рис. 3C). Это открытие контрастирует с условиями без стресса, в которых РНК осадка и общая РНК показали лишь небольшие различия. Таким образом, стрессовый осадок демонстрирует состав РНК, отличный от состава общей РНК во время арсенитного стресса, вероятно, из-за образования стрессовых гранул во время арсенитного стресса и высвобождения мРНК из полисом.
Рис. 3. Транскриптомы осадка гранул
стрессированной РНК воспроизводимы.(A) Трехмерная диаграмма рассеяния, изображающая нормализованное количество считываний из библиотек РНК-seq из трех экземпляров осадка гранул подвергнутых стрессу РНК. (B) Трехмерная диаграмма рассеяния, изображающая нормализованное количество считываний из библиотек РНК-seq из трех экземпляров полной РНК, подвергшихся стрессу. (C) Таблица, изображающая парные коэффициенты корреляции Пирсона между осадками гранул РНК и общей РНК в стрессированных клетках. (D) График МА, отображающий значения log 2 кратного изменения (осадок RG под стрессом / общая РНК под стрессом) в зависимости от численности (FPKM). Гены обозначены цветом в зависимости от их значения.Значимые гены ( P <0,01) окрашены в красный цвет, а незначимые ( P > 0,01) — в синий. (E) Диаграмма рассеяния стрессированного обогащения RG, полученного из фибробластов мыши, по сравнению с стрессированным обогащением RG в клетках U-2 OS. (F) Онтология генов для обогащенных и истощенных транскриптов.
Мы выполнили дифференциальный анализ обогащения, чтобы идентифицировать транскрипты, которые обогащены и истощены из осадка подвергнутой стрессу РНК. Анализ дифференциального обогащения выявил многочисленные транскрипты, которые были значительно ( P <0.01), обогащенного ( n = 1053) и истощенного ( n = 1449) осадка подвергнутых стрессу гранул РНК (рис. 3D). Показатели обогащения из осадка гранул РНК, подвергнутого стрессу, положительно коррелировали с осадком гранул, подвергнутых ранее стрессу, выделенных из фибробластов мыши ( R = 0,35) (рис. 3E), что согласуется с этими двумя экспериментами, дающими аналогичные результаты. Онтология генов для обогащенных транскриптов показала, что РНК, которые были обогащены в осадке, обычно кодируют транскрипты, обычно участвующие в развитии или биогенезе (рис.3F). В частности, гены, участвующие в нейрогенезе, были по-разному обогащены в осадке, что согласуется с тем фактом, что SG-обогащенные транскрипты имеют тенденцию обнаруживать большое количество перекрывающихся длинных транскриптов, которые важны для нормального развития мозга (13).
Мы наблюдали с помощью генной онтологии, что транскрипты, участвующие в метаболизме, были значительно истощены из осадка подвергнутого стрессу RG (рис. 3F). Это прямо противоположно условиям без стресса, в которых транскрипты, кодирующие метаболические транскрипты, были обогащены в осадке.Мы связываем это изменение с тем фактом, что арсенитный стресс вызывает глобальное отключение трансляции, и только меньшая часть генов продолжает трансляцию. Поскольку метаболические гены, скорее всего, больше не будут транслироваться, они не будут связаны с полисомами и, следовательно, не будут находиться в состоянии RNP, достаточно тяжелом для осаждения при 16000 × г . Это может объяснить, почему метаболические гены смещаются во фракцию цитозольного супернатанта в стрессовых условиях.
Мы также сравнили, насколько близко подвергнутый стрессу осадок РНК напоминал осадок нестрессированной РНК.Мы наблюдали значительную корреляцию между осадком подвергнутой стрессу РНК и осадком нестрессированной РНК ( R = 0,76) (рис. 4A). Однако между этими двумя условиями была существенная разница. Чтобы определить показатели, которые могут объяснить различия в обогащении, мы изучили, как различные особенности РНК коррелируют с дифференциальным обогащением.
Рис. 4
Характеристика осадка гранул подвергнутой стрессу РНК. (A) Диаграмма рассеяния, изображающая корреляцию между обогащением напряженных гранул по сравнению с ненагруженным обогащением гранул.(B) Графики в виде прямоугольников, показывающие длины транскриптов для обогащенных гранулами и обедненных гранулами транскриптов во время стресса от арсенита и в условиях без стресса. (C) То же, что и панель B, но для значений эффективности трансляции (TE) (получено из Subtelny et al. [18]). (D) То же, что и панель B, но имеет цветную маркировку в зависимости от длины стенограммы. (E) То же, что и панель B, но с цветовой кодировкой по значениям TE.
Длина транскрипта коррелировала с обогащением осадка как в стрессовых, так и в нестрессированных условиях (рис. 4B). Однако отметим, что в напряженных условиях зависимость обогащения от длины была более выраженной, чем в ненапряженных.Этот вывод согласуется с предыдущим наблюдением, что длина играет важную роль в определении того, какие РНК обогащены стрессовыми гранулами (8, 14), и согласуется с тем, что стрессовые гранулы обогащаются в осадке при 16000 × г (8, 10, 15–17).
Мы также исследовали, как эффективность трансляции коррелирует с локализацией РНК. Во время арсенитного стресса осажденные РНК имели значительно более низкие значения ТЕ (18), чем РНК, которые были истощены из осадка (рис. 4С).Это открытие контрастирует с условиями без стресса, во время которых транскрипты с более высокими значениями TE осаждаются. Таким образом, плохо транслируемые транскрипты имеют тенденцию осаждаться во время арсенитного стресса, тогда как транскрипты с более высокими значениями ТЕ имеют тенденцию осаждаться в условиях отсутствия стресса.
Поскольку длина и трансляция коррелируют с обогащением гранул РНК во время арсенитового стресса, мы исследовали, объясняют ли эти два параметра различия между обогащением гранул без стресса и обогащением гранул в стрессовом состоянии.Цветовая кодировка диаграммы разброса обогащения осадка в стрессовом состоянии по сравнению с обогащением осадка без стресса по длине транскрипта показала, что транскрипты, обогащенные осадком во время стресса, имеют тенденцию быть длиннее, чем транскрипты, обогащенные осадком в условиях отсутствия стресса (рис. 4D). Мы также наблюдали аналогичную зависимость для эффективности трансляции, при которой плохо транслируемые транскрипты были обогащены в подвергнутой стрессу осадке, в то время как эффективно транслируемые транскрипты были обогащены в нестрессированной осадке (рис.4E). В соответствии с этими результатами, с помощью многомерного линейного регрессионного анализа, длина и эффективность трансляции составляют дополнительные 20% дисперсии между обогащением гранул без стресса и с учетом стресса (данные не показаны).
Взятые вместе, эти результаты показывают, что существует общая корреляция между РНК, которые осаждаются в стрессовых и нестрессированных условиях. Кроме того, различия в корреляции между обогащением гранул в стрессовых и нестрессированных условиях можно в значительной степени объяснить, учитывая дополнительные факторы длины транскрипта и эффективности трансляции.В стрессовых условиях более длинные плохо транслируемые транскрипты имеют тенденцию накапливаться в осадке, аналогично тому, что можно было бы ожидать в стрессовых гранулах (8), в то время как в условиях стресса относительно более короткие и более эффективно транслируемые транскрипты также накапливаются в осадке RG.
Сравнение транскриптомов RG с транскриптомом стрессовых гранул. Ключевой нерешенный вопрос заключается в том, насколько хорошо транскриптомы гранул РНК сравниваются с транскриптомами стрессовых гранул, основанных на более селективной иммунной очистке, и можно ли использовать простое гранулирование в качестве подходящего заместителя для полный протокол очистки стрессовых гранул.В то время как Namkoong et al. сравнили транскриптом осадка стрессовых гранул из фибробластов мыши с нашим предыдущим транскриптомом стрессовых гранул из клеток U-2 OS ( R 2 = 0,49), мы хотели использовать аналогичный подход в той же клеточной линии U-2 OS. Таким образом, мы повторно проанализировали данные в нашем предыдущем транскриптоме стрессовых гранул (8), используя тот же вычислительный конвейер, который мы использовали для транскриптомов гранул РНК (рис. 5A).
FIG 5
Полисомы, вероятно, объясняют различия в стрессовом транскриптоме RG и в транскриптоме стрессовых гранул.(A) График МА, отображающий значения log 2 -кратного изменения (РНК стрессовых гранул / стрессовая общая РНК) в зависимости от численности (FPKM). Гены обозначены цветом в зависимости от их значения. Значимые гены окрашены в красный цвет ( P <0,01), а незначимые гены окрашены в синий цвет ( P > 0,01). (B) Диаграмма разброса обогащения напряженных гранул по сравнению с обогащением напряженных гранул. (C) Диаграмма разброса обогащения напряженных гранул по сравнению с обогащением напряженных гранул. (D) То же, что и панель C, но с цветовой кодировкой по обогащению гранул в ненапряженных условиях.(E) Значения R2 из множественного линейного регрессионного анализа для обогащения в стрессовых гранулах по сравнению с обогащением в виде SG или обогащения в стрессовых гранулах в сравнении с обогащением в SG и обогащении в стрессовых гранулах. (F) То же, что и панель C, но с цветовой кодировкой в соответствии с процентным содержанием GC. (G) То же, что и панель C, но с цветовой кодировкой по доле оптимальных кодонов на транскрипт. (H) Анализ TapeStation осадка RG во время стресса и SG РНК с использованием дополнительных этапов иммуноочистки.
Сначала мы сравнили транскриптом стрессовой гранулы с транскриптомом нестрессированной гранулы РНК.Мы наблюдали, что оценки обогащения стрессовых гранул и оценки обогащения стрессовых гранул не коррелировали (фиг. 5B). Таким образом, транскриптом стрессовых гранул не просто представляет собой РНК, которые имеют тенденцию к осаждению в отсутствие стресса.
Напротив, оценки обогащения стрессовых гранул и оценки обогащения стрессовых гранул РНК показали значительную положительную корреляцию (фиг. 5C). Это демонстрирует, что осадок гранул стресса повторяет транскриптом гранул стресса до определенной степени, что согласуется с идеей, что гранулы РНК являются разумным заместителем для приготовления гранул полного стресса.
Полисомы могут загрязнять транскриптом RG. Было неясно, почему стрессированный транскриптом RG не был более точным отражением транскриптома SG, определенного с помощью иммуноочистки. При значении R 2 , равном всего 0,32, это означает, что две трети дисперсии оценок обогащения стрессовых гранул по сравнению с оценками обогащения стрессовых RG остаются необъясненными. Отчасти эта вариабельность может заключаться в несоответствии между циклами секвенирования и эффектами партии. По нашим оценкам, до 20% дисперсии в оценках обогащения связаны с артефактами секвенирования, основанными на различиях между повторностями секвенирования (см. Значения R 2 на рис.1А и 2А). Однако это означает, что ~ 40% дисперсии остается необъяснимой.
Одна возможность состоит в том, что РНК, которые имеют более высокую склонность к осаждению как в стрессовых, так и в нестрессовых условиях, являются источником неспецифического загрязнения в методе очистки RG-гранул. Если бы это предсказание было верным, мы бы ожидали, что эти загрязняющие РНК также будут иметь высокую склонность к осаждению даже в нестрессовых условиях. Поэтому мы закодировали нашу диаграмму рассеяния обогащения стрессовых гранул по сравнению с обогащением стрессовых гранул за счет обогащения осадка РНК в нестрессовых условиях.Поразительно, что мы наблюдали, что транскрипты, которые имеют тенденцию обогащаться гранулами стрессированного RG по сравнению с гранулами стресса, являются теми же транскриптами, которые имеют тенденцию осаждаться в условиях отсутствия стресса (рис. 5D). Взятые вместе, эти данные также предполагают, что осадок гранул подвергнутой стрессу РНК содержит дополнительные РНК, которые обогащаются даже в отсутствие стресса.
Чтобы исследовать, какая разница между обогащением стрессовых гранул и стрессовых гранул может быть объяснена загрязнением РНК, которые осаждаются независимо от стрессовых условий, мы провели многомерный регрессионный анализ методом наименьших квадратов.Этот анализ позволяет количественно изучить, какая часть дисперсии данного распределения объясняется комбинацией нескольких переменных. Когда мы добавили оценки обогащения гранул без стресса RG в качестве дополнительной независимой переменной, мы наблюдали значительно улучшенный показатель R 2 , равный 0,536, по сравнению с нашим исходным показателем R 2 , равным 0,32 (рис. 5E). Это говорит о том, что гранулы подвергнутой стрессу РНК содержат некоторые загрязняющие РНК, которые обогащаются даже в отсутствие стресса.
Мы дополнительно проанализировали данные, чтобы определить, почему некоторые РНК осаждаются как в стрессовых, так и в нестрессовых условиях. Мы наблюдали, что транскрипты, которые предпочтительно обогащаются осадком стрессированного RG по сравнению с гранулами стресса, имеют тенденцию иметь более высокое содержание GC, в то время как транскрипты, которые предпочтительно обогащаются стрессовыми гранулами по сравнению с осадком стрессированного RG, имеют тенденцию иметь более низкое содержание GC (рис. 5F).
Поскольку систематическая ошибка содержания GC может влиять на использование кодонов, мы предположили, что использование кодонов играет роль в различении относительного обогащения транскрипта в подвергнутом стрессу осадке RG от стрессовых гранул.Чтобы проверить эту гипотезу, мы составили список неоптимальных и оптимальных кодонов и сравнили процент оптимальных кодонов в каждом транскрипте. Мы наблюдали явное смещение, при котором транскрипты, содержащие более высокие фракции оптимальных кодонов, предпочтительно обогащались фракцией стрессированных RG по сравнению с стрессовыми гранулами (рис. 5G). Мы интерпретируем эти результаты как потенциально представляющие загрязнение полисомами в осадке RG, подвергающемся стрессу. Состав кодонов коррелирует с трансляцией: транскрипты с большим процентом оптимальных кодонов, как правило, показывают более высокие скорости трансляции.В соответствии с полисомами, загрязняющими осадок RG, с помощью анализа TapeStation мы наблюдали сильные полосы рРНК при гранулировании, в то время как при очистке SG мы видим более слабые рибосомные полосы, сопровождаемые более темным мазком в тех размерах, где проходят длинные РНК (от 1,5 до 6 т.п.н.) (Рис. 5H). Таким образом, мы предполагаем, что транскрипты, которые продолжают транслироваться во время стресса, до некоторой степени загрязняют подвергнутый стрессу осадок RG, поскольку очень большие полисомы и стрессовые гранулы имеют тенденцию иметь сходные седиментационные свойства.
Интересное наблюдение состоит в том, что существует популяция РНК, которая обогащена как стрессированным транскриптомом RG, так и транскриптомом SG, а также обогащена нестрессированным транскриптомом RG (Fig. 5D). Таким образом, этот набор РНК обогащен SG, но также склонен к осаждению даже в отсутствие стресса. Одна возможность состоит в том, что это представляет собой популяцию РНК, которые могут быть собраны в более мелкие стрессовые гранулы даже в отсутствие стресса и, таким образом, будут склонны к образованию осадка и все же будут обогащены стрессовыми гранулами.
Анализ транскриптомов Р-тельцов и их отношения к стрессовым гранулам и гранулам РНК. Эукариотические клетки также содержат Р-тельца, которые также должны оседать в условиях, которые генерируют транскриптомы RG (3, 16). Таким образом, мы проанализировали, насколько близко ненапряженный или подвергнутый стрессу осадок напоминал транскриптом Р-тельцов, что было ранее определено путем сбора Р-тельцов на сортировщике клеток в условиях отсутствия стресса из клеток HEK293 (7).
Мы наблюдали, что нестрессированные и стрессированные транскриптомы осадка РНК практически не коррелировали с транскриптомом Р-телец (рис.6А и В). Это согласуется с нашим анализом, описанным выше, предполагающим, что осадок RG без стресса представляет собой гены, кодирующие метаболические ферменты и транскрипты, нацеленные на митохондрии, а осадок RG в стрессовом состоянии представляет собой смесь RNPs стрессовых гранул и загрязняющих транскриптов, которые осаждаются независимо от стресса. Это говорит о том, что осаждения РНК недостаточно для очистки РНК, которые связываются с Р-тельцами, и что необходима дополнительная очистка.
Рис. 6.
Сравнение транскриптомов SG и P-телец.(A) Диаграмма рассеяния, изображающая обогащение РНК в ненапряженном осадке RG по сравнению с Р-тельцами. (B) Диаграмма рассеяния, изображающая обогащение РНК в подвергнутом стрессу осадке RG по сравнению с Р-тельцами. (C) Диаграмма рассеяния, изображающая обогащение РНК Р-тельцами по сравнению со стрессовыми гранулами. (D) То же, что и панель C, но с цветовой кодировкой в соответствии с содержимым GC.
Сравнение стрессовой гранулы и транскриптома Р-тельца. Мы хотели изучить, как обогащение Р-тельца коррелирует с оценками обогащения стрессовых гранул, о которых мы ранее сообщали (8).Таким образом, мы повторно проанализировали исходный транскриптом P-body (7) с помощью того же вычислительного конвейера, в котором мы анализировали транскриптом стрессовых гранул. Мы наблюдали, что сравнение транскриптома P-телец с транскриптомом стрессовых гранул дало относительно сильную положительную корреляцию ( R = 0,53) (рис. 6C). Эта корреляция заслуживает внимания, особенно потому, что мы сравниваем транскриптом P-тела из ненапряженного состояния с вызванными стрессом гранулами стресса в разных типах клеток.Это предполагает, что некоторые из тех же правил, которые играют роль в разделении РНК на стрессовые гранулы, также влияют на разделение РНК на Р-тельца. В соответствии с таким сходством плохая трансляция и увеличенная длина коррелируют с накоплением мРНК как в P-тельцах, так и в SG (см. Ниже) (7, 8).
РНК, которые обогащаются P-тельцами, имеют сильное смещение AU (12). Учитывая эту предвзятость, мы хотели изучить степень, в которой нуклеотидный состав коррелирует со степенью, в которой транскрипты преимущественно обогащаются PB по сравнению со стрессовыми гранулами.Транскрипты, которые были сильно обогащены PB, но не обогащены SG, как правило, имели более высокие уровни состава AU, тогда как транскрипты, которые были сильно обогащены в SG, но не обогащены в PB, имели тенденцию иметь более высокие уровни содержания GC (рис. 6D). Изучение содержания AU с помощью многомерного линейного регрессионного анализа дало дополнительные 11% дисперсии (данные не показаны). Таким образом, содержимое AU может играть роль в определении того, какие транскрипты предпочтительно делятся на P-тельца, а не на стрессовые гранулы.
Транскриптом Р-тельца смещается во время стресса. Есть по крайней мере две возможности относительно того, почему содержание AU модулирует локализацию РНК в Р-теле. Во-первых, транскрипты, обогащенные Р-тельцами, обычно плохо транслируются и имеют высокий уровень неоптимальных кодонов (12). Таким образом, состав AU может определять обогащение РНК Р-тельцами из-за плохой трансляции. Во-вторых, белки P-тельца могут иметь более сильное сродство к AU-богатым транскриптам, чем GC-богатые транскрипты, что приводит к преимущественному обогащению AU-богатых транскриптов в PB.
Чтобы различить эти две модели, мы исследовали транскриптом P-body во время арсенитного стресса, в котором трансляция ограничена. Если обогащенные Р-тела транскрипты разделяются на Р-тела посредством их ассоциации с РНК-связывающими белками Р-телец, можно ожидать, что нестрессированный транскриптом Р-телец будет демонстрировать высокую степень корреляции с подвергнутыми стрессу арсенитом Р-тельцами. транскриптом. Однако, если транскрипты в основном локализуются в P-теле из-за их нетранслирующего статуса, можно ожидать, что транскриптом P-телец будет изменяться во время стрессовых состояний.Обоснование, лежащее в основе этой гипотезы, состоит в том, что во время стресса трансляция значительно снижается; таким образом, доступный пул субстратов РНК должен значительно увеличиться и включать больше GC-богатых транскриптов.
Чтобы определить, какие РНК локализуются в Р-тельцах во время арсенитного стресса, мы очистили Р-тельца от обработанных арсенитом клеток U-2 OS. Мы адаптировали нашу схему очистки стресс-гранул для очистки P-тел. Вкратце, Р-тельца выделяли посредством последовательных циклов центрифугирования, после чего Р-тельца дополнительно очищали посредством иммунопреципитации с помощью антитела против EDC3.Кроме того, мы также иммуноочистили этим методом Р-тельца из клеток G3BP1Δ / G3BP2Δ, которые не могут образовывать стрессовые гранулы при обработке арсенитом (19, 20). Мы добавили этот контроль, поскольку SG могут стыковаться с Р-тельцами (2), и их стыковка повысила вероятность того, что иммуноочищенные препараты Р-тельца будут иметь некоторое загрязнение стрессовыми гранулами.
Мы наблюдали, что библиотеки, полученные из клеток дикого типа (WT) и G3BP1 Δ / G3BP2 Δ после очистки P-телец, дали воспроизводимые результаты, которые заметно отличаются от результатов для общего цитоплазматического транскриптома (рис.7A и B). Транскриптомы PB зависели от антитела Edc3, поскольку при использовании контрольного антитела IgG для стадии иммуноочистки РНК не выделялась (фиг. 7C и D). Более того, транскриптомы Р-телец WT и G3BP1 Δ / G3BP2 Δ были очень похожи, что позволяет предположить, что стрессовые гранулы не являются существенным загрязнителем в наших препаратах Р-телец (рис. 7E). Анализ транскриптома Р-тельца из клеток дикого типа во время стресса выявил следующие моменты.
FIG 7
Сравнение стрессированного и нестрессированного транскриптома Р-тельца.(A) Таблица, показывающая попарные значения r 2 между PB РНК и общей РНК в клетках, подвергнутых стрессу арсенита. (B) То же, что и панель A, но для клеток с нокаутом G3BP1 / G3BP2 (ko). (C) Анализ TapeStation для контроля пониженного уровня IgG. (D) Анализ TapeStation на EDC3 РНК Р-телец. (E) Диаграмма рассеяния, изображающая корреляцию между обогащением Р-телец дикого типа и обогащением Р-телец G3BP1Δ / G3BP2Δ во время стресса. (F) Обогащение Р-тел в условиях без стресса по сравнению с обогащением Р-тел во время арсенитного стресса.(G) Обогащение стрессовых гранул по сравнению с обогащением Р-телами во время арсенитного стресса.
Во-первых, когда мы сравнили стрессированный транскриптом P-тела с транскриптомом исходного нестрессированного транскриптома P-тела, мы наблюдали довольно сильную положительную корреляцию ( R = 0,55) (рис. 7F). Однако стрессированный транскриптом Р-тельца показал большее сходство с транскриптомом стрессовой гранулы ( R = 0,85) (рис.7G), чем ненапряженный транскриптом Р-тельца в условиях стресса, индуцированного арсенитом ( R = 0.55) (Рис. 4F). Это указывает на то, что транскриптом Р-тельца смещается во время стресса из-за более широкого доступного пула нетранслирующих мРНК.
Когда мы исследовали параметры, которые коррелируют с обогащением P-тела, мы видим заметный сдвиг от нестрессовых к стрессовым условиям. В частности, нестрессированные обогащенные транскрипты Р-телец имели тенденцию демонстрировать более высокие уровни неоптимальных кодонов, чем подвергнутые стрессу обогащенные транскрипты Р-телец (фиг. 8A-C). Это открытие согласуется с нашей гипотезой о том, что трансляция определяет, какие субстраты доступны для секвестрации Р-тел.Более того, в стрессовых условиях транскриптом P-тел показывает значительно более сильную корреляцию с длиной транскрипта (рис. 8D-F) и меньшую зависимость от эффективности трансляции. Это указывает на то, что длина играет большую роль в определении состава Р-телец в условиях арсенитного стресса, когда большинство мРНК сильно репрессируются трансляцией.
FIG 8
Взаимодействие между трансляцией и длиной определяет транскриптом P-body. (A) Диаграмма рассеяния, показывающая корреляцию между долей оптимальных кодонов и обогащением P-тел в нестрессовых условиях.(B) То же, что и панель A, но при арсенитном напряжении. (C) График в виде прямоугольника, показывающий долю оптимальных кодонов для обогащенных и истощенных транскриптов в стрессированных и нестрессированных Р-телах. (D) Диаграмма рассеяния, показывающая корреляцию между длиной и обогащением P-тел в условиях отсутствия стресса. (E) Диаграмма рассеяния, показывающая корреляцию между длиной и обогащением P-тел в стрессовых условиях. (F) График в виде прямоугольников, показывающий длину транскрипта для обогащенных и истощенных транскриптов в стрессированных и нестрессированных Р-телах.
Анализ, описанный выше, предложил специфические мРНК, которые предпочтительно были бы обогащены Р-тельцами, и что эти мРНК должны увеличивать свое разделение на Р-тельца во время стресса.Чтобы проверить это предсказание, мы исследовали, как обогащенные транскрипты из наших данных RNA-seq коррелировали с обогащением P-телец посредством одномолекулярной флуоресцентной гибридизации in situ (smFISH). Сначала мы исследовали локализацию SPEN, который, как ранее было показано, обогащен Р-тельцами в нестрессированных клетках (7). В соответствии с этим наблюдением мы обнаружили, что около 12,5% молекул SPEN проявляли ассоциацию с Р-тельцами в условиях отсутствия стресса (рис. 9А). Мы также наблюдали, что SPEN был сильно обогащен стрессированным транскриптомом P-телец.В соответствии с этим наблюдением около 40% транскриптов SPEN обнаруживают ассоциацию с Р-тельцами в клетках, обработанных арсенитом (фиг. 9B). Таким образом, SPEN ассоциируется с Р-телами в условиях отсутствия стресса, но дополнительно усиливает свою ассоциацию с PB во время арсенитного стресса.
FIG 9
smFISH-валидация транскриптома P-тела. (A) smFISH SPEN и IF RCK, маркер P-тела, в условиях отсутствия стресса. (B) То же, что и панель A, но при арсенитном напряжении. (C) smFISH AHNAK и IF RCK, маркер P-тела, в условиях отсутствия стресса.(D) То же, что и панель C, но при арсенитном напряжении. (E) smFISH DYNC1h2 и IF RCK, маркера P-тела, в условиях отсутствия стресса. (F) То же, что и панель E, но при арсенитном напряжении. (G) smFISH из GAPDH и IF из RCK, маркера P-тела, в условиях отсутствия стресса. (H) То же, что и панель G, но при арсенитном напряжении. (I) Количественная оценка процента транскриптов, связанных с PBs, с помощью smFISH для SPEN, AHNAK, DYNC1h2 и GAPDH в отсутствие стресса и арсенитного стресса. (J) Диаграмма рассеяния, изображающая корреляцию между обогащением Р-тел, оцененным с помощью smFISH, и обогащением Р-тел, оцененным с помощью RNA-seq.
Мы также исследовали некоторые транскрипты, которые ранее не были показаны как обогащенные нестрессированным транскриптомом Р-телец, но были высоко обогащены стрессированным транскриптомом Р-телец, такие как AHNAK и DYNC1h2. В соответствии с анализом РНК-seq, эти транскрипты показали ограниченную ассоциацию с Р-тельцами в отсутствие стресса (фиг. 9C и E), но показали повышенные уровни ассоциации во время арсенитного стресса (фиг. 9D и F). Это наблюдение подтвердило, что состав РНК PB может изменяться в стрессовых условиях.
Наконец, мы исследовали глицеральдегид-3-фосфатдегидрогеназу (GAPDH) в качестве контроля, так как она была истощена как из стрессовых, так и из стрессированных Р-тел с помощью последовательности РНК. В соответствии с этим наблюдением, GAPDH показал ограниченную связь с Р-тельцами в присутствии или в отсутствие стресса (рис. 9G-I). Т.о., P-тельца не просто увеличивают ассоциацию со всеми РНК во время стресса, но, вероятно, следуют правилам, сходным с таковыми для SGs (Fig. 9I).
Количественное определение локализации мРНК в PB выявило сильную корреляцию между smFISH и RNA-seq, предполагая, что наше секвенирование достоверно и отражает реальность in vivo РНК, ассоциированных с P-тельцами (рис.9J). Это наблюдение подтвердило, что ассоциация РНК с Р-тельцами может сильно изменяться во время стресса и что во время стресса состав РНК PB аналогичен составу SG.
ОБСУЖДЕНИЕ
Были описаны два различных метода очистки и анализа транскриптомов SG. В одном методе РНК, обогащенные тяжелым осадком в стрессовых условиях, обозначаются как транскриптом RG и, как предполагается, коррелируют с транскриптомом SG (10). В качестве альтернативы SG можно обогатить дифференциальным центрифугированием, а затем иммуноочистить на последующей стадии, метод, который дает транскриптом SG, строго подтвержденный smFISH.Здесь мы напрямую сравнили эти два метода.
Наше сравнение показывает, что хотя стрессированный транскриптом RG показывает некоторое сходство с транскриптомом SG, он больше похож на транскриптом RG из нестрессированных клеток (рис. 4 и 5). Мы наблюдаем, что РНК, обогащенные тяжелым осадком, также включают митохондриальные транскрипты и некоторые хорошо транслируемые РНК, которые не обогащены SG. Более того, ряд РНК обогащается в осадке как в стрессовых, так и в стрессовых условиях, выявляя фоновую популяцию загрязняющих РНК при использовании осадка RG для определения транскриптома SG.Таким образом, осадок RG под стрессом является приближением транскриптома SG, но более точное представление определяется иммуноочисткой стрессовых гранул (8).
Наши результаты показывают, что состав РНК Р-тел резко отличается в стрессовых и стрессовых условиях, когда Р-тела увеличиваются в размере и количестве (2, 3). Это основано на нашем анализе транскриптома Р-тельца во время арсенитного стресса в клетках ОС U-2 по сравнению с транскриптомом Р-тельца из нестрессированных клеток HEK293 (7).Наш анализ показал изменение свойств РНК, которые обогащаются Р-тельцами в стрессовых и нестрессовых условиях. В отсутствие стресса мРНК, обогащенные P-тельцами, сильно коррелируют с показателями плохо транслируемых мРНК (рис. 8). Это согласуется с тем, что Р-тельца накапливают нетранслирующие мРНК, что свидетельствует о том, что состав Р-тельцов в отсутствие стресса в первую очередь диктуется РНК, которые плохо транслируются (12). Напротив, во время стресса, когда трансляция большинства мРНК сильно ингибируется, мы наблюдали, что обогащение мРНК в Р-тельцах больше не зависит от показателей, связанных с плохой трансляцией, и вместо этого смещается в сторону более длинных РНК (рис.8).
Одним из ограничений этого анализа является то, что мы сравниваем транскриптом PB между двумя разными клеточными линиями. Мы провели это сравнение, потому что популяция PB в нестрессированных клетках U-2 OS довольно мала, и было непрактично очищать PB из этой линии клеток в нестрессовых условиях. Тем не менее, поскольку наши эксперименты с smFISH (рис. 9) демонстрируют явное обогащение специфических мРНК в PB при стрессе, мы считаем, что это сравнение является, по крайней мере, разумным приближением.
Основываясь на этом сдвиге в составе P-тела, мы предполагаем, что состав P-тела определяется различными характеристиками в стрессовых и нестрессовых условиях (рис.10), где низкая эффективность трансляции является основной детерминантой разделения P-тел в нестрессовых условиях, но в стрессовых условиях, когда большинство мРНК больше не транслируются, мРНК дифференцированно разделяются на PB в зависимости от длины и, возможно, других показателей, таких как способность мРНК связываться с ключевыми компонентами P-тела, такими как Ddx6, Pat1l, Edc3 и Edc4.
FIG 10
Трансляция определяет состав РНК Р-телец. Модель, изображающая критическую роль трансляции в определении состава РНК PB.(Слева) В безударных условиях для секвестрации P-тела доступны только расшифровки с плохими метриками перевода. (Справа) В условиях стресса, в которых трансляция ограничена, состав РНК PB изменяется, чтобы включать больше РНК и больше походить на транскриптом SG.
Мы также заметили, что состав Р-тел и стрессовых гранул был очень похож во время стресса (рис. 7). Это сходство не связано с загрязнением SG наших препаратов P-телец, поскольку мы наблюдаем подобный транскриптом PB даже в клетках G3BP1 Δ / G3BP2 Δ , которые не собираются для образования SG (рис.7). Это согласуется с ситуацией во время стресса, когда большинство мРНК вышли из трансляции и теперь доступны для сборки либо в SG, либо в PB. Сходство мРНК внутри PBs и SG во время стресса согласуется с предыдущими экспериментами FISH, показывающими, что мРНК от одного и того же гена могут быть обнаружены как в SG, так и в PB (2), в наших экспериментах smFISH (рис.9) и экспериментах с одной молекулой. показывая, что отдельные мРНК могут проходить между PB и SG (21). Это предполагает, что специфические особенности отдельных мРНК, кодируемых одним и тем же геном, определяют их ассоциацию с PB или SG.Например, одна возможность состоит в том, что мРНК с более коротким поли (A) хвостом, который может способствовать связыванию комплекса PB, Pat1 / Lsm1-7 (22), будут молекулами, обогащенными PB, в то время как молекулы с более длинным поли ( A) «хвосты» предпочтительно накапливаются в SG. С другой стороны, модификации РНК могут изменить связывание РНК-связывающих белков, чтобы повлиять на их разделение (23). Дальнейшая работа по определению того, как отдельные молекулы мРНК делятся на PB и SG, будет важна для решения этих проблем.
МАТЕРИАЛЫ И МЕТОДЫ
Условия выращивания ОС U-2 и реагенты.Клетки OS остеосаркомы человека U-2 поддерживали в среде Игла, модифицированной Дульбекко (DMEM) с высоким содержанием глюкозы, 10% фетальной бычьей сыворотки и 1% пенициллин-стрептомицин при 37 ° C и 5% CO 2 и использовали во всех экспериментах. .
Выделение осадка RG и P-телец. Клетки OS U-2 выращивали до ~ 75% слияния в трех 15-сантиметровых чашках для культивирования. За час до стресса среду для культивирования клеток заменяли свежей средой. Затем клетки обрабатывали 0,5 мМ NaAsO 2 в течение 1 ч при 37 ° C и 5% CO 2 .После стресса клетки промывали один раз средой, переносили в пробирки Falcon и осаждали при 1500 × г в течение 3 мин. После аспирации среды гранулы немедленно замораживали в жидком N2 и хранили при -80 ° C до выделения осадка гранул РНК.
Протокол выделения гранул РНК был основан на нашей оригинальной схеме очистки стрессовых гранул, за исключением шагов иммунопреципитации (8, 9). Вкратце, осадок оттаивали на льду в течение 5 мин, ресуспендировали в 1 мл буфера для лизиса SG (50 мМ трис-HCl [pH 7.4], 100 мМ ацетат калия, 2 мМ ацетат магния, 0,5 мМ дитиотреитол, 50 мкг / мл гепарина, 0,5% NP-40, полный ингибитор протеазы, не содержащий мини-ЭДТА [11836170001; Sigma-Aldrich], 1 ед. / Мкл РНКазина плюс ингибитор РНКазы [N2615; Promega]) и пропустили 7 раз через иглу 25 размера 5/8, прикрепленную к шприцу на 1 мл. Лизаты центрифугировали при 1000 × г в течение 5 минут при 4 ° C для осаждения остатков клеток. Пятьдесят микролитров супернатантов переносили в новые микроцентрифужные пробирки для выделения общей РНК, а остаток использовали для остальной части протокола.
Следующие шаги были выполнены для выделения осадка RG млекопитающих и экстракции его РНК. (i) 950 мкл супернатанта центрифугировали при 16000 × г в течение 20 минут при 4 ° C для осаждения ядер SG. (ii) Полученный супернатант отбрасывали, и осадок ресуспендировали в 1 мл буфера для лизиса SG. (iii) Шаги i и ii были повторены. (iv) Полученный осадок затем ресуспендировали в 300 мкл буфера для лизиса SG и центрифугировали при 850 × г в течение 2 минут при 4 ° C. (v) Супернатант, представляющий собой осадок РНК, переносили в новую пробирку.
Для выделения общей РНК и осадочной РНК добавляли реагент RTrizolLS (10296-028; Thermo Fisher Scientific) и экстрагировали РНК в соответствии с протоколом производителя. После осаждения изопропанолом осадок РНК ресуспендировали в 50 мкл свободных от РНКаз H 2 O.
P-тельца очищали, как описано для SG (8), за исключением того, что использовали 20 мкл антитела против Edc3 (ab168851). чтобы изолировать Р-тела.
Стрессовые условия. Для изучения локализации мРНК при стрессе были выполнены следующие стрессовые условия.Для экспериментов с арсенитным стрессом клетки обрабатывали 0,5 мМ арсенита натрия (S7400; Sigma-Aldrich) в течение 1 часа. Клетки фиксировали после стресса 4% параформальдегидом (NC0179595; Fisher Scientific).
Построение библиотеки и секвенирование РНК. Качество РНК оценивали с помощью анализа TapeStation. Библиотеки кДНК с парными концами получали в центре секвенирования института Biofrontiers с использованием KAPA HyperPrep с RiboErase. Библиотеки кДНК секвенировали на устройстве с высоким выходом NextSeq (150 циклов [2 × 75]).
Анализ данных секвенирования. Качество чтения оценивалось с помощью FastQC. Адаптеры Illumina были обрезаны с использованием Trimmomatic 0.32 в парном режиме и режиме PE (24). Индексный геном был получен из GENCODE (выпуск 19 GRCh47.p13). Чтения были выровнены с использованием TopHat (версия 2.0.6) и Bowtie2 (версия 2.0.2) и следующих параметров: −b2 −fast −microexon-search (25, 26). Анализ дифференциальной экспрессии был выполнен с использованием Cuffdiff (версия 2.2.1) с параметрами по умолчанию (27). Анализ онтологии генов выполнялся с использованием инструментов Консорциума генных онтологий (http: // www.geneontology.org/). Длины транскриптов были получены от Ensembl’s Biomart Tool.
smFISH analysis.smFISH выполняли, как описано ранее (8). Для окрашивания P-тел мы использовали иммунофлуоресценцию (IF) против RCK, маркера PB (MDL PD009).
Доступность данных. Последовательность операций доступна через NCBI GEO (GSE138988).
Калькулятор напряжений
Этот калькулятор напряжений поможет вам решить задачи механики, связанные с напряжением, деформацией и модулем Юнга. За несколько простых шагов вы узнаете зависимость напряжения от деформации для любого материала, который остается эластичным.Мы также научим вас рассчитывать деформацию и применять уравнение напряжения.
Как рассчитать деформацию и напряжение
Деформация определяется как мера деформации — пропорция между изменением длины и исходной длиной объекта. Например, если взять резинку и растянуть ее так, чтобы она была вдвое длиннее, чем изначально, то деформация будет равна 1 (100%).
Формула деформации:
ε = ΔL / L₁ = (L₂ - L₁) / L₁
L₁ обозначает начальную длину, L₂ — конечную длину, а ΔL — изменение длины.Обратите внимание, что деформация безразмерна.
Напряжение, с другой стороны, является мерой давления, которое частицы материала оказывают друг на друга. Он определяется как сила, действующая на объект на единицу площади. Однако это отличается от давления; при расчете напряжения рассматриваемая площадь должна быть настолько маленькой, чтобы анализируемые частицы считались однородными. Если брать во внимание большую площадь, расчетное напряжение обычно является средним значением.
Уравнение напряжения:
σ = F / A
F обозначает силу, действующую на тело, а A обозначает площадь.Единицы измерения напряжения такие же, как и единицы давления — Паскали (символ: Па) или Ньютоны на квадратный метр.
Положительное напряжение означает, что объект находится в напряжении — он «хочет» удлиниться. Отрицательное напряжение означает, что оно находится в состоянии сжатия и «хочет» стать короче.
Модуль Юнга (напряжение относительно деформации)
Если материал линейно упругий , то напряжение и деформация напрямую связаны следующей формулой:
E = σ / ε
E — это модуль упругости или модуль Юнга .Это материальная константа, разная для каждого вещества.
Что такое линейно-упругое поведение материала? Если мы прикладываем напряжение к материалу, деформация увеличивается пропорционально. Это может быть верно только для некоторого диапазона напряжений — после достижения определенного значения материал может сломаться или деформироваться. Под текучестью понимается увеличение деформации при постоянном напряженном состоянии.
Пример расчетов
Предположим, мы хотим найти модуль Юнга стали. Для этого мы подготовили стальной стержень, который тянули с большим усилием.
- Мы решили, что сила тяги штанги будет равна 30 кН (
30 * 10³ Н). - Определяем размеры стержня. Предположим, что длина составляет 2 м (2000 мм), а площадь поперечного сечения — 1 см² (
1 * 10⁻⁴ м²). - Мы заметили, что стержень удлинился на 3 мм.
- Рассчитываем деформацию стержня по формуле:
ε = ΔL / L₁ = 3/2000 = 0,0015. - Рассчитываем напряжение по формуле напряжений:
σ = F / A = 30 * 10³ / (1 * 10⁻⁴) = 300 * 10⁶ = 300 МПа. - Наконец, разделим напряжение на деформацию, чтобы найти модуль Юнга стали:
E = σ / ε = 300 * 10⁶ / 0,0015 = 200 * 10⁹ = 200 ГПа.
Модуль упругости, ед.
Единицы модуля Юнга такие же, как единицы давления и напряжения: Паскали или ньютоны на квадратный метр. В единицах СИ
1 Па = 1 Н / 1 м² = 1 кг · м / с² / м² = 1 кг / (м · с²)
Если вас интересует механика, попробуйте калькулятор крутящего момента.
Белки Р-тельца регулируют перестройку транскрипции для обеспечения устойчивости к стрессу репликации ДНК
Удаление
LSM1 в значительной степени ремоделирует транскриптом
LSM1 необходим для регуляции функционального образования Р-телец, а также для деградации или стабильности конкретных мишеней мРНК 16,17,18 . Чтобы идентифицировать набор мРНК-мишеней регуляции Р-тельца, мы использовали RNA-seq для профилирования транскриптомов клеток дикого типа (WT) и lsm1∆ клеток в присутствии и в отсутствие стресса репликации, индуцирующего лекарственную гидроксимочевину. (HU) (Рис.1а). Эксперимент проводили в двух экземплярах на общей РНК, лишенной рРНК, и дифференциальную экспрессию оценивали с использованием протокола Tuxedo 19 . Гены, показывающие дифференциальную экспрессию со статистическим значением P менее 0,05 после коррекции Бенджамини-Хохберга 19 , в нашем анализе рассматривались как совпадения. На транскриптом сильно повлияла делеция LSM1 (рис. 1b, c; дополнительные данные 1). Как и ожидалось, учитывая роль Lsm1 в деградации РНК 16,17,18 , мы обнаружили, что количество 333 мРНК увеличивалось при удалении LSM1 .Неожиданно подобное количество мРНК (258) уменьшилось в изобилии (рис. 1b, c; дополнительные данные 1). Хотя мы не определили, являются ли эти мРНК прямыми мишенями для P-телец, возможно, что ассоциация мРНК с Lsm1 в некоторых случаях является защитной и предотвращает зависимую от экзосом деградацию 20 . Альтернативно, отсутствие LSM1 может стабилизировать репрессоры транскрипции, что косвенно приводит к снижению количества мРНК, как это наблюдалось в клетках, лишенных экзонуклеазы 5ʹ – 3 РНК Xrn1 21 .
Рис. 1
Транскриптом в значительной степени реконструируется, когда функциональные Р-тельца отсутствуют. a Обзор стратегии RNA-seq, используемой для измерения содержания РНК в lsm1 ∆ клеток. b Отношения log2 экспрессии генов, нанесенные в виде тепловой карты. Отношения экспрессии, сравнивающие обработку HU с необработанным состоянием (+ HU: -HU), у дикого типа ( WT ) и lsm1 ∆, нанесены на график слева . Отношения экспрессии для сравнения lsm1 ∆ клеток с WT-клетками во время нормального роста и воздействия HU ( lsm1 ∆: WT) нанесены на график справа . c На графике нанесено число дифференциально экспрессируемых транскриптов в lsm1 ∆ во время репликационного стресса. d Распределение соотношений экспрессии lsm1 ∆: WT отображается в виде прямоугольных диаграмм для всех генов ( серый ) и дифференциально экспрессируемых генов ( белый ). P -значения взяты из непарных тестов Вилкоксона. e Сводные данные по обогащению взаимодействия для мРНК с увеличенной численностью в lsm1 ∆ ( желтый ) и со сниженной численностью в lsm1 ∆ ( blue ) во время нормального роста (без лекарств) и во время репликационного стресса (HU) . P -значения были рассчитаны с использованием биномиального теста. f сети обогащения термина GO для мРНК, которые дифференциально экспрессировались в lsm1 ∆ во время нормального роста, для мРНК с повышенной численностью ( желтый ) и мРНК с пониженной численностью ( синий ). Узлы закрашены в соответствии со значениями P ( самых темных красных , обозначающих наименьшее значение P ). Размер узла указывает частоту, с которой данный член GO встречается в S.cerevisiae геном. Кромки соединяют очень похожие термины GO, и толщина кромки представляет степень сходства. г Сети обогащения терминов GO для мРНК, которые дифференциально экспрессировались в lsm1 ∆ во время лечения HU (гены HU-core)
В клетках WT обработка HU вызвала значительное изменение количества мРНК. Сотни транскриптов показали увеличение или уменьшение численности всего через 1 час обработки HU (дополнительный рис.1 и дополнительные данные 1). После 4 часов обработки HU 1134 и 1178 транскриптов показали увеличение или уменьшение количества соответственно (дополнительный рисунок 1). Мы обнаружили разумную положительную корреляцию между нашими данными и предыдущими транскриптомными исследованиями микроматрицы в HU 6 или метилметансульфонате (MMS) 7, 22 (дополнительный рис. 2a, b). Корреляция с данными, полученными с использованием отдельного агента стресса репликации, MMS, указывает на то, что значительная часть идентифицированной нами программы транскрипции связана со стрессом репликации ДНК (дополнительный рис.2б). Некоторые изменения в экспрессии генов, которые мы наблюдаем, вероятно, являются результатом синхронизации клеточного цикла в ранней S-фазе, которая происходит при добавлении HU. И WT, и lsm1∆ подвергаются сходным арестам HU, однако, и поэтому эффекты клеточного цикла вряд ли будут влиять на идентификацию дифференциально экспрессируемых генов при сравнении WT и lsm1 ∆ (дополнительный рис. 3).
Делеция LSM1 оказала драматическое влияние на количество мРНК во время лечения HU.От 499 (через 1 час после обработки HU) до 1203 транскриптов (через 4 часа после обработки HU) увеличилось количество в штамме lsm1 ∆ (рис. 1b, c и дополнительные данные 1) по сравнению с контролем дикого типа. Неожиданное снижение количества транскриптов, которое мы наблюдали в отсутствие обработки HU в lsm1 ∆ было более значительным в присутствии HU, где количество мРНК уменьшилось от 322 до 1051. Почти половина дифференциально экспрессируемых генов в клетках lsm1 ∆ была подавлена, что снова указывает на непредвиденную положительную или защитную роль белков Р-телец в регуляции экспрессии генов (рис.1b, c и дополнительные данные 1). В целом, однако, распределение соотношений обилия мРНК для дифференциально экспрессируемых генов показало статистически подтвержденное положительное смещение, указывая на то, что стабилизирующий эффект на РНК, наблюдаемый при делеции LSM1 , сильнее, чем неожиданный дестабилизирующий эффект (рис. 1d). Интересно, что большая часть (до 53%) транскриптов, на количество которых повлияла обработка HU и делеция LSM1 , также были затронуты обработкой MMS, что позволяет предположить, что Lsm1-регулируемые транскрипты меняются в количестве во время стресса репликации ДНК в целом. (Дополнительный рис.2в).
Наконец, чтобы подтвердить, что дифференциально экспрессируемые гены, которые мы идентифицировали, не зависели от используемого метода анализа данных, мы применили два дополнительных анализа для идентификации дифференциально экспрессируемых генов: EBSeq 23 и edgeR 24 . От 34 до 79% генов, идентифицированных в нашем первоначальном анализе, также были идентифицированы с помощью EBSeq или edgeR, в зависимости от анализируемой временной точки (дополнительная таблица 1).
Дифференциально экспрессируемые гены функционально обогащены
Анализ существующих баз данных 25 выявил умеренное обогащение как генетических, так и физических взаимодействий в дифференциально экспрессируемых генах в lsm1 ∆, как во время нормального роста, так и при стрессе репликации ДНК (рис.1д). Эти данные показывают, что гены, которые дифференциально экспрессируются в lsm1 ∆ по сравнению с WT, с большей вероятностью имеют общие функциональные биологические связи, и это верно как во время невозмущенного роста, так и при наличии репликационного стресса.
Далее мы оценили функциональные связи между дифференциально экспрессируемыми генами путем тестирования на обогащение терминов биологического процесса онтологии генов (GO). Среди генов с увеличенным содержанием мРНК в lsm1∆ мы отметили обогащение членами GO терминов, связанных с синтезом АТФ, особенно генов, кодирующих компоненты цепи переноса электронов, предполагая роль Р-телец в регуляции энергетических транзакций во время нормального клеточного роста (рис. .1f и дополнительные данные 2). Помимо метаболизма рибонуклеотидов, небольшое дополнительное функциональное обогащение наблюдалось среди генов со сниженной экспрессией в lsm1 ∆, предполагая, что количество мРНК снижается в отсутствие функциональных Р-телец во время нормального роста клеток может быть в значительной степени неспецифичным.
Чтобы оценить функциональное обогащение в присутствии репликационного стресса, мы определили основной набор HU-чувствительных генов как гены, дифференциально экспрессируемые только при воздействии HU и в двух последовательных временных точках (601 увеличение численности и 417 снижение численности).Кроме того, мы удалили гены, которые, как известно, участвуют в общей реакции на стресс окружающей среды, из нашего основного набора генов, чувствительных к HU (72 гена в наборе повышенной численности (42 гена активированы в ответе на стресс окружающей среды) и 49 генов в наборе пониженной численности. (35 снижена в реакции на стресс окружающей среды)) 26 . Гены, участвующие в нескольких аспектах функции митохондрий, в частности, в импорте белка в митохондрии, были обогащены набором генов, количество мРНК которых увеличилось в HU в lsm1∆ , как и гены, кодирующие субъединицы протеасомы (рис.1g и дополнительные данные 2). Гены, кодирующие регуляторы клеточного цикла, были обогащены генами, количество мРНК которых снизилось в HU в клетках lsm1∆ (рис. 1g и дополнительные данные 2). Функциональное обогащение предполагает, что Р-тельца подавляют митохондриальный биогенез и деградацию белков и активируют гены, участвующие в контроле развития клеточного цикла во время стресса репликации, индуцированного HU.
Таким образом, в дополнение к основному регуляторному эффекту, который функциональные Р-тела оказывают на экспрессию генов в невозмущенных клетках, мы определяем дополнительный уровень регуляции обилия транскриптов белками Р-телец, как положительных, так и отрицательных, что проявляется при репликации стресс присутствует.По сравнению с невозмущенными клетками белки Р-тельца регулируют различные клеточные процессы во время воздействия стресса репликации, что согласуется с конкретной биологической ролью Р-тельцов в ответе на стресс репликации ДНК.
Супрессоры чувствительности к HU мутантов P-телец
Установив, что белок P-телец Lsm1 прямо или косвенно регулирует количество мРНК во время репликационного стресса, мы попытались идентифицировать мРНК, подавление регуляции которых на P-телец важен для репликации стрессоустойчивости.Мы пришли к выводу, что чувствительность к HU мутантов P-тельца обусловлена увеличением количества критических мРНК-мишеней P-тельца. Следовательно, делеция генов, кодирующих эти мишени, должна подавлять чувствительность HU мутантов P-тельца (Fig. 2a). Поскольку LSM1 и PAT1 имеют большинство генетических взаимодействий любых двух компонентов P-тела 27 , мы провели подавляющие экраны как lsm1∆ , так и pat1∆ (рис. 2b и дополнительные данные 3), используя метод синтетического генетического массива (SGA) 28 для систематической инактивации 4971 гена (~ 76% генома) на фоне lsm1 ∆ или pat1 ∆.Мы оценили приспособленность каждого двойного мутанта, измеряя и сравнивая размер колонии в присутствии и в отсутствие HU в трех экземплярах. Значения пригодности были нормализованы для удаления артефактов пластины и / или закрепления и преобразованы в баллы Z . Распределение баллов Z было сходным для фонов деформации lsm1 ∆ и pat1 ∆ (рис. 2b). Двойные мутанты с оценкой Z больше +1 считались супрессорами чувствительности к HU. Поскольку отдельные подавляющие экраны имели низкий уровень истинно положительных результатов (~ 27%; 3 кандидата проверены, после восстановления деформации, из 11 случайно выбранных попаданий подавителя), мы ограничили наш анализ подавителями, идентифицированными как в lsm1, ∆ и pat1 ∆ экраны.В общей сложности 71 ген при инактивации был способен подавлять чувствительность как lsm1 ∆, так и pat1 ∆ HU. Среди этих генов-мишеней Р-тельцов 11 также имеют повышенное количество транскриптов в отсутствие LSM1 в 2 или более раз подряд после обработки HU (рис. 2c). Независимая реконструкция двойных мутантов pat1 ∆ и lsm1 ∆ с каждым из 11 генов привела к валидации 6 предполагаемых генов-мишеней: ARL3 , ACF4 , HHT1 , TMA1 , RRS и YOX1 и повышенное содержание мРНК в HU для 5 из этих транскриптов было подтверждено двумя независимыми методами анализа данных. ACF4 был подтвержден edgeR, но не EBSeq (дополнительная таблица 2). Мутации в каждом из них подавляли чувствительность как lsm1 ∆, так и pat1 ∆ HU (рис. 2d). Ни в одном случае подавление не было полным, что позволяет предположить, что несколько целей P-body вносят вклад в приспособленность во время репликационной реакции на стресс. Биологическая функция 6 генов описана на рис. 2e. Два гена ( RRS1 и TMA16 ) кодируют ассоциированные с рибосомами белки 29, 30 , что позволяет предположить, что подавление способности к трансляции белков важно для устойчивости к стрессу репликации ДНК.Два ( ACF4 и YOX1 ) кодируют мишени фосфорилирования циклинзависимой киназы Cdc28 31 . Важно отметить, что среди шести идентифицированных нами генов один ( HHT1) , как известно, является мишенью для P-тел во время репликационного стресса 32 . Таким образом, наша стратегия идентифицировала пять новых генов, экспрессия которых токсична во время репликационного стресса, когда функция P-тельца нарушена.
Рис. 2
Комбинирование систематического скрининга супрессоров с данными обилия мРНК для идентификации мишеней Р-тельцов при репликационном стрессе. a Обзор экрана для определения супрессоров чувствительности HU мутантов P-тельца. b
Z — нормализованные значения пригодности HU для двойных мутантов lsm1 ∆ xxx и pat1 ∆ xxx . Гены, которые были идентифицированы на обоих экранах, обозначены зеленым . Подтвержденные совпадения указаны справа . c Обилие мРНК во время репликационного стресса отображается в виде тепловой карты для одиннадцати генов-кандидатов на пересечении результатов скрининга супрессоров и данных о содержании мРНК.Отношение ∆: WT log2 lsm1 нанесено на график для указанных моментов времени. d Анализ чувствительности HU для 6 проверенных генов-кандидатов среди 11 генов, идентифицированных в c . Десятикратные разведения ночной культуры наносили на среду YPD с HU или без нее. e Биологические функции проверенных генов-кандидатов
YOX1 регуляция с помощью P-тел критична во время стресса HU
Yox1 является гомеобоксным репрессором транскрипции, который регулирует экспрессию регулируемых клеточного цикла и генов репликации ДНК 33 .Вместе с Mcm1, Yox1 связывается с промоторами генов, регуляция которых является критической во время перехода M / G1 клеточного цикла 33 . Инактивация белка Yox1 важна во время репликационного стресса у Schizosaccharomyces pombe
34 , а сверхэкспрессия YOX1 вызывает резкое увеличение потери хромосом у S. cerevisiae .
35 . Стресс репликации ДНК заставляет Yox1 повторно локализоваться из ядра в цитоплазму 8 , подтверждая, что экспрессия мишеней Yox1 может играть важную роль в ответе на стресс репликации.Учитывая его связь с ответом на репликационный стресс, мы спросили, регулируется ли экспрессия YOX1 белками Р-тельца.
Мы впервые подтвердили, что уровни мРНК YOX1 увеличиваются у мутантов P-тельца. Когда мы исследовали мРНК YOX1 с помощью количественной ПЦР с обратной транскрипцией (ОТ-ПЦР), мы обнаружили, что в lsm1 ∆ и pat1 ∆ YOX1 накапливалась мРНК относительно клеток WT (дополнительный рисунок 4). Удаление PAT1 отменяет образование Р-телец во время репликационного стресса 8 , в то время как потеря LSM1 приводит к накоплению нефункциональных Р-телец 18 , что позволяет предположить, что деградация YOX1 в некоторой степени зависит от образования функциональных Р-телец во время репликационного стресса.Однако, поскольку Lsm1-зависимая и Pat1-зависимая деградация мРНК также может происходить в отсутствие видимых гранул P-телец, возможно, что регулирование мРНК YOX1 белками P-телец может происходить на сайтах, внешних по отношению к видимым гранулам P-телец.
Затем мы проверили, снижает ли инактивация YOX1 чувствительность мутантов P-телец к различным стрессам или является ли она специфичной для стресса репликации ДНК, вызванного обработкой HU (рис. 3). Удаление LSM1
36 или PAT1 придает чувствительность к борной кислоте, однако делеция YOX1 не спасла ни одну из чувствительности (рис.3а). Мутант lsm1∆ также чувствителен к тепловому стрессу 37 , а температурную чувствительность не спасает yox1∆ (рис. 3а). Ген YOX1 имеет паралог, YHP1 , поэтому мы также оценили эффект удаления YHP1 на стрессоустойчивость в lsm1∆ и pat1∆ (рис. 3а). Делеция YHP1 мало влияла на HU, тепло или чувствительность к борной кислоте мутантов P-тельца. Кроме того, мРНК YHP1 не подвергалась повышенной регуляции в lsm1 ∆ (дополнительные данные 1).Мы пришли к выводу, что делеция YOX1 спасает чувствительность к HU мутантов P-тел, играя небольшую роль в других стрессовых реакциях P-тел, и что паралог YHP1 вряд ли будет релевантной мишенью для P-тел во время стресса репликации.
Фиг.3
YOX1 Токсичность специфична для стресса репликации ДНК. a Серийные 10-кратные разведения указанных штаммов наносили на YPD с или без HU, борной кислоты (BA) или при высокой температуре (40 ° C).Планшеты были визуализированы через 2–3 дня роста, чтобы оценить чувствительность каждого штамма к каждой обработке. b Серийные 5-кратные разведения WT (BY4741), lsm1 ∆ или pat1 ∆ клеток, содержащих pBY011 (пустой вектор) или pBY011- YOX1 ( GAL1-10pr-YOX1 ) синтетических плазмид. полная среда без урацила, содержащая рафинозу или рафинозу и галактозу, с HU или без нее. Планшеты были визуализированы через 7 дней роста, чтобы оценить влияние экспрессии YOX1 на чувствительность к HU
Мы пришли к выводу, что если P-телец-зависимая деградация мРНК YOX1 является критической для устойчивости к репликационному стрессу, то избыточная экспрессия YOX1 должна повышать чувствительность мутантных штаммов P-телец к HU.В самом деле, экспрессия YOX1 с промотора GAL1-10 была токсичной как в фонах lsm1 ∆, так и pat1 ∆ (рис. 3b). Вместе эти данные подтверждают вывод о том, что негативная регуляция экспрессии YOX1 белками Р-тельца важна конкретно для устойчивости к HU-стрессу.
Транскрипт
YOX1 локализуется в P-тельцах
Чтобы оценить, вероятно ли, что негативная регуляция экспрессии YOX1 происходила непосредственно в цитоплазматических гранулах P-телец, которые образуются во время репликационного стресса, мы отслеживали местоположение YOX1 мРНК в живых клетках с использованием системы визуализации мРНК mTAG 38 .Мы ввели повторы MS2 в 3ʹ UTR гена YOX1 и экспрессировали белок оболочки MS2, слитый с зеленым флуоресцентным белком (GFP), для визуализации мРНК YOX1 . Р-тельца визуализировали одновременно путем совместной экспрессии слитого белка Pat1-mCherry. Система тегов MS2 может поставить под угрозу визуализацию полноразмерных мРНК, способствуя Xrn1-зависимому накоплению продуктов деградации мРНК 39, 40 , поэтому мы исследовали локализацию мРНК YOX1 как в WT, так и в клетках, удаленных для XRN1 .Кроме того, мы пришли к выводу, что, поскольку транскрипт YOX1 потенциально деградирует на Р-тельцах, где Xrn1 является преобладающей 5ʹ – 3ʹ экзорибонуклеазой 41 , делеция XRN1 должна облегчить визуализацию мРНК YOX1 за счет ее стабилизации.
YOX1 мРНК образует гранулы в ~ 6% WT-клеток как в необработанных условиях, так и после обработки HU (рис. 4a, b). Делеция XRN1 резко увеличивала образование гранул мРНК YOX1 и Р-телец 18 в необработанных условиях и после обработки HU (рис.4а, б). Контрольные штаммы, экспрессирующие белок оболочки MS2-GFP, но лишенные повторов MS2 в 3ʹUTR YOX1 , показали очень мало гранул (рис. 4b), демонстрируя, что для формирования фокуса MS2-GFP требовалось YOX1 , помеченное повторами MS2. Важно отметить, что гранулы YOX1 совместно локализованы с маркером P-тел Pat1 как в клетках WT, так и в клетках xrn1 ∆ в нормальных условиях и после обработки HU (рис. 4a, b). Из 1092 исследовано гранул YOX1 , 25,8 ± 8.8% мРНК YOX1 и гранул Pat1 локализованы совместно в xrn1 ∆ необработанных клетках. Сходные уровни совместной локализации были обнаружены с другими транскриптами и маркером P-телец Edc3 42 . Обработка HU приводила к появлению крупных агрегатов в красном канале, что затрудняло количественную оценку совместной локализации гранул мРНК YOX1 с Pat1-mCherry (рис. 4a). Тем не менее, когда наблюдались дискретные фокусы Pat1, мы могли обнаружить совместную локализацию с гранулами мРНК YOX1 (рис.4а). Интересно, что накопление гранул мРНК YOX1 , которое очевидно при делеции XRN1 , коррелирует с повышенной чувствительностью к HU (рис. 4c) и увеличением количества транскриптов YOX1 ( xrn1 ∆: дикий тип = 3,7 ± 1,8). Как и в случае с lsm1 ∆, чувствительность HU xrn1 ∆ восстанавливается за счет делеции YOX1 (рис. 4c), что указывает на то, что мРНК YOX1 является критическим транскриптом для чувствительности HU xrn1 ∆ .Таким образом, неспособность расщепить мРНК YOX1 либо на уровне образования функциональных Р-телец, либо на уровне нуклеолитической деградации мРНК, приводит к чувствительности к HU. Эти данные подтверждают мнение о том, что контроль обилия мРНК YOX1 важен для ответа на стресс репликации.
Фиг.4
YOX1 мРНК образует цитоплазматические гранулы и накапливается в Р-тельцах. a Репрезентативные изображения клеток, коэкспрессирующих MS2-CP-GFP и Pat1-mCherry, с YOX1 , помеченными повторами MS2, или без них, до и после 2 часов обработки HU.Цитоплазматические гранулы оценивали как в WT, так и в клетках xrn1 ∆. Шкала составляет 7 мкм. Стрелки указывают на гранул мРНК YOX1 , которые локализуются совместно с гранулами Pat1-mCherry. b Количественное определение гранул мРНК YOX1 в необработанных и обработанных HU клетках. Среднее количество очагов на клетку наносится на график для каждой из не менее трех повторов. Сплошная полоса указывает среднее значение повторов, P -значения взяты из тестов Стьюдента t , а n указывает общее количество исследованных клеток. c Серийные 10-кратные разведения указанных штаммов наносили на YPD с или без HU. Планшеты были визуализированы через 3–4 дня роста, чтобы проверить подавление чувствительности yox1 ∆HU путем делеции XRN1
В совокупности эти данные показывают, что мРНК YOX1 локализуются в Р-тельцах как при нормальном росте, так и во время репликационного стресса, вызванного HU, что согласуется с моделью, в которой количество мРНК YOX1 регулируется белками Р-тельца, включая Xrn1, прямо у P-body.
Yox1 накапливается в ядре в
lsm1 ∆ клеток
Yox1 и его паралог Yhp1 связываются с транскрипционным антирепрессором Mcm1, что приводит к репрессии мишеней Mcm1 33 . Установив, что транскрипт YOX1 увеличивается в клетках lsm1 ∆ в присутствии HU (рис. 2c), мы проверили, увеличивается ли также содержание белка Yox1. Yox1 накапливается в цитоплазме после обработки HU 8 , поэтому мы исследовали содержание белка Yox1-GFP в отдельных клетках и в отдельных ядрах как в клетках WT, так и в lsm1 ∆ клеток (рис.5). Клетки, лишенные LSM1 , имели повышенные уровни Yox1-GFP в ядре и в цитоплазме (рис. 5c, d), что согласуется с увеличением содержания мРНК YOX1 в lsm1∆ . Повышенный уровень Yox1-GFP в ядре в lsm1∆ сохранялся после обработки HU не менее 2 ч (рис. 5г). Иммуноблот-анализ Flag-tagged Yox1 подтвердил увеличение белка Yox1 в lsm1 ∆ клеток (рис. 5b). Стоит отметить, что изобилие белка Yox1 не точно отражает изобилие мРНК YOX1 , что предполагает дополнительную регуляцию на уровне трансляции и / или стабильности белка.Эти данные указывают на то, что в ядре больше Yox1, когда мРНК YOX1 не деградирует эффективно на P-тельцах, и предполагают, что гены-мишени Yox1 будут экспрессироваться на более низких уровнях, когда функция P-тельца нарушена.
Рис. 5
Репрессор Yox1 более распространен в клетках, лишенных LSM1. a Типичные изображения клеток, экспрессирующих Hta2-mCherry и Yox1-GFP до и после 2 часов обработки HU. Масштабные линейки имеют размер 5 мкм, и показаны контуры ячеек. b Анализ уровня белка Yox1-Flag с помощью иммуноблоттинга до и после лечения HU. Pgk1 использовался в качестве контроля загрузки. Маркеры молекулярной массы показаны на необрезанном изображении на дополнительном рис. 5. Показан один типичный эксперимент. Нормализованная интенсивность клеточной ( c ) или ядерной ( d ) флуоресценции Yox1-GFP в клетках WT и lsm1 ∆. Каждая точка представляет данные для отдельной реплики указанного штамма (для 8 технических реплик для каждой из 2 биологических реплик).Сплошная полоса указывает среднее значение повторов, а P -значения из теста Стьюдента t указаны
Сеть мишеней Yox1 важна для устойчивости к HU
Чтобы исследовать последствия повышенной экспрессии YOX1 во время репликационного стресса, мы использовали RNA-seq для измерения количества транскриптов во время сверхэкспрессии YOX1 в присутствии HU (рис. 6а). , b и дополнительные данные 4).Экспрессия от 96 (через 2 часа) до 156 генов (через 4 часа) была снижена в штамме с избыточной экспрессией YOX1 . От одной трети (2 часа) до половины (4 часа) генов, которые подавлялись при сверхэкспрессии YOX1 , также подавлялись в клетках ∆ lsm1, что согласуется с модуляцией функции Yox1 с помощью Lsm1 (рис. 6b, c) . Этот эффект статистически подтверждается тем, что lsm1 ∆ генов с пониженной регуляцией были сильно обогащены среди генов с пониженной регуляцией, идентифицированных при сверхэкспрессии YOX1 (рис.6в). Интересно, что ряд генов с измененной экспрессией в присутствии HU и избыточной экспрессии YOX1 не были известны Yox1 мишенями 43 (фиг. 6b, фиолетовая панель ). Следовательно, потенциальный репертуар регуляторных мишеней Yox1 расширяется во время репликационного стресса.
Рис. 6
Избыток Yox1 репрессирует гены, которые имеют решающее значение для приспособленности клеток во время репликационного стресса. a Обзор эксперимента RNA-seq. Клетки WT, содержащие векторы pBY011 (пустой вектор) или pBY011- YOX1 ( GAL1-10pr-YOX1 ), культивировали в синтетической среде, содержащей рафинозу.Сверхэкспрессия YOX1 индуцировалась добавлением галактозы, а стресс репликации индуцировался обработкой HU. РНК экстрагировали в указанные моменты времени. b Тепловая карта соотношений экспрессии, созданная с помощью иерархической кластеризации, показывающая гены, которые дифференциально экспрессировались при сверхэкспрессии YOX1 . Представлены два различных соотношения: + HU: -HU сравнивает значения экспрессии в контрольном штамме (пустой вектор) между состоянием HU и необработанным состоянием, а GAL1-10pr-YOX1 : пустой вектор сравнивает значения экспрессии между YOX1 -выполняющий штамм и контрольный штамм в каждый момент времени. Голубые столбики указывают на гены, которые подавлялись через 2 часа и / или 4 часа в HU в lsm1 ∆ по сравнению с WT (как на рис. 1). Фиолетовые столбики обозначают гены, которые описаны как мишени Yox1 в базе данных Yeastract. c Число генов со сниженным содержанием мРНК при сверхэкспрессии YOX1 , с перекрытием генов, которые также имеют пониженное содержание мРНК в lsm1 ∆, представлено в виде столбчатой диаграммы. Указанные значения P были рассчитаны с использованием гипергеометрического теста. d Идентификация потенциальных мишеней Yox1 во время стресса репликации на пересечении трех наборов данных. Тепловые карты представлены в b . e Данные пригодности в HU для одиночных мутантов, идентифицированных в d . Каждая точка данных представляет собой одну реплику ( n = 5). Горизонтальные полосы представляют собой среднее значение повторений. P -значения взяты из теста студента t . f Зависимая от состояния отрицательная генетическая сеть взаимодействий между 8 целями в d . Толщина красной кромки пропорциональна разнице между пригодностью двойного мутанта в отсутствие и в присутствии 25 мМ HU
Удивительно, но программа репрессии транскрипции YOX1 , которую мы идентифицировали, не была обогащена генами, индивидуально участвующими в устойчивости к HU (гипергеометрический P = 0,992 через 2 часа и P = 0,998 через 4 часа). Мы сосредоточились на 8 мишенях Yox1, которые также были подавлены в лсм1 ∆ и вызвали синтетическую болезнь HU в лсм1 ∆ (рис.6d), полагая, что наиболее критические мишени Yox1 будут затронуты сверхэкспрессией Yox1, делецией LSM1 и усугубят чувствительность HU на lsm1 ∆. Мы отметили, что только два ( ALD6 и RAD54 ) были чувствительны к стрессу репликации ДНК сами по себе (рис. 6e). Чтобы проверить возможность того, что избыточность может объяснить относительное отсутствие чувствительности к единичным мутантам HU, мы сконструировали 20 комбинаций мутантов с двойным нокаутом по 8 мишеням (за исключением RTC1 , которые не были доступны в нашей коллекции делеций).Мы проанализировали приспособленность, отслеживая рост в жидкой среде в присутствии или в отсутствие HU. Используя этот подход, мы идентифицировали 7 негативных генетических взаимодействий, которые усугублялись наличием стресса репликации ДНК (рис. 6f, дополнительные данные 5). В трех взаимодействиях участвовали гены, которые не были чувствительны к HU как одиночные мутанты, а в четырех — RAD54 . Следовательно, функциональная избыточность среди генов в регуломе YOX1 может маскировать роли отдельных генов в устойчивости к стрессу репликации.Мы предполагаем, что подробный анализ двойного мутантного пространства в регуломе YOX1 выявит множество дополнительных взаимодействий, которые придают чувствительность HU.
Вместе эти данные предполагают, что накопление Yox1, которое происходит в отсутствие функционального пути P-телец, приводит к подавлению регуляции транскрипционной сети, которая в целом поддерживает приспособленность клеток во время стресса репликации.
Детоксикация ацетальдегида имеет решающее значение при воздействии HU
Хотя транскрипционная программа YOX1 в целом, вероятно, вносит свой вклад в устойчивость к стрессу репликации, мы также проверили, можем ли мы определить роли отдельных генов-мишеней.8 идентифицированных мишеней подавляются при сверхэкспрессии YOX1 и когда функция P-телец нарушена (фиг. 6d). Поэтому мы экспрессировали каждый ген из промотора GAL1-10 в штамме lsm1∆ в присутствии HU, чтобы проверить, может ли какой-либо ген индивидуально восстановить чувствительность HU lsm1 ∆ (рис. 7a, b). Экспрессия ICS2 или ALD6 увеличивала приспособленность lsm1∆ во время репликационного стресса (рис. 7a, b), указывая на то, что репрессия ICS2 и ALD6 посредством зависимой от белка P-тела деградации YOX1 мРНК способствует устойчивости к стрессу репликации.Мы идентифицировали сайты связывания как для Yox1, так и для его корепрессора Mcm1 в промоторных областях длиной 1000 пар оснований ALD6 и ICS2 , используя YeTFaSCo 44 (дополнительная таблица 3), хотя также возможно, что оба являются косвенными мишенями. Наконец, lsm1∆ чувствителен к ацетальдегиду (рис. 7c), как и следовало ожидать, если экспрессия альдегиддегидрогеназы, кодируемой ALD6 , снижается, когда P-тельца не функционируют.
Рис. 7
Дерепрессия мишени Yox1 ALD6 имеет решающее значение для детоксикации ацетальдегида и повышения устойчивости к стрессу репликации ДНК. a Относительный рост lsm1 ∆ клеток, сверхэкспрессирующих указанные гены, по сравнению с lsm1 ∆ клеток, содержащих контрольный вектор. Каждая точка данных представляет собой одну реплику. Каждый эксперимент повторяли не менее трех раз и каждый раз включали в себя две независимые биологические повторы. Сплошная полоса показывает среднее значение. b Кривые роста lsm1 ∆ клеток, сверхэкспрессирующих ALD6 или ICS2 ( красный ) или lsm1 ∆ клеток, содержащих контрольный вектор ( черный ) в присутствии HU. c Серийные 10-кратные разведения указанных штаммов наносили на YPD с ацетальдегидом или без него. Чашки были визуализированы через 3-4 дня роста. d
Слева : Тепловая карта значений пригодности клеток, выращенных в присутствии ацетальдегида, гидроксимочевины или комбинации обоих препаратов. Значения пригодности относятся к необработанному состоянию для трех независимых повторов. Справа : тепловая карта Synergy. Указанные значения соответствуют разнице между наблюдаемой пригодностью для данной комбинации лекарственных средств и ожидаемой пригодностью. e Пригодность клеток, выращенных в присутствии ацетальдегида, гидроксимочевины или комбинации обоих препаратов в указанных концентрациях. Каждая точка данных представляет собой одну реплику. P -значения взяты из теста студента t . f Интенсивности Rnr3-GFP у указанных мутантов относительно WT. Каждая точка данных представляет собой одну реплику. P -значения взяты из теста студента t . г Типичные микроскопические изображения указанных штаммов, экспрессирующих Rnr3-GFP и RPL39pr-TdTomato. Шкала : 5 мкм. ч Развитие клеточного цикла штамма WT и ald6 ∆ после блока альфа-фактора и высвобождения в YPD, содержащем 200 мМ HU
Мы исследовали механизм, с помощью которого ALD6 способствует устойчивости к стрессу репликации. Сначала мы использовали количественную ОТ-ПЦР, чтобы подтвердить, что количество мРНК ALD6 уменьшается в lsm1∆ (дополнительный рис. 6). Ald6 превращает ацетальдегид в ацетат, и, таким образом, клетки ald6 ∆ накапливают большее количество ацетальдегида 45 .Ацетальдегид индуцирует межцепочечные сшивки ДНК, которые блокируют репликацию и транскрипцию ДНК 46 и, следовательно, потенциально могут быть токсичными для клеток, испытывающих стресс репликации ДНК. Мы обнаружили, что HU и ацетальдегид обладают синергическим действием в диапазоне концентраций, что указывает на то, что присутствие ацетальдегида во время стресса репликации, вызванного HU, действительно токсично (рис. 7d, e). Удаление ALD6 вызывает сильную конститутивную активацию контрольной точки S-фазы, о чем свидетельствует индукция экспрессии RNR3 в клетках ald6 ∆ до того же уровня, что и в rad54∆ , одном из самых сильных индукторов зависимых от контрольных точек . Экспрессия RNR3 15 (рис.7е, г). В соответствии с накоплением повреждений ДНК, блокирующих репликацию, в клетках, лишенных Ald6, и синергизмом между ацетальдегидом и HU, мы обнаружили, что ald6 ∆ клеток демонстрируют медленное продвижение S-фазы в HU по сравнению с клетками WT.
Вместе эти данные предполагают, что накопление ацетальдегида вызывает эндогенный стресс репликации, который сенсибилизирует клетки, испытывающие дополнительные источники стресса репликации ДНК. Следовательно, дерепрессия ALD6 во время стресса репликации, вызванного HU, способствует приспособленности за счет удаления ацетальдегида, уменьшения повреждения ДНК и уменьшения барьеров для синтеза ДНК.
Влияние психосоциального стресса на дофаминергическую функцию и реакцию на острый стресс
Существенные изменения:
Помимо рассмотрения всех вопросов, поднятых рецензентами (см. Ниже), авторы должны уделять особое внимание следующим темам:
1) Объясните и обоснуйте, почему стресс-задача (MIST) была выполнена за два часа до ПЭТ-сканирования.
См. Наш ответ рецензенту 2 (ниже).
2) Как можно разделить острые контрольные и экспериментальные эффекты и как разрешить взаимодействие с хроническими невзгодами?
Мы рассмотрели комментарии рецензентов по этому поводу ниже.Мы прокомментировали это в Обсуждении:
«Тем не менее, потенциальное ограничение нашей комбинации факторов стресса на раннем этапе развития и стресса взрослого человека состоит в том, что он может смешивать эффекты в раннем периоде жизни (т.е. вероятное программирование) с острыми стрессорами в более позднем возрасте, которые возникают после периодов чувствительности к развитию. […] Поэтому необходима дальнейшая работа, чтобы разобраться в влиянии острых и хронических стрессоров на дофаминовую систему ».
3) Разделы, касающиеся фундаментальной науки и активности популяции дофамина, нуждаются в пояснении.
Мы прояснили эти разделы по просьбе рецензентов.
Рецензент №1:
В этой рукописи авторы исследуют влияние предшествующего воздействия психосоциального стресса на показатели функции DA у людей. Статья довольно интересная и потенциально важная; однако есть некоторые проблемы, которые необходимо решить:
1) Связь с литературой по животным в том виде, в каком она написана, сбивает с толку в отношении высвобождения DA и стресса по сравнению со способностью синтеза у людей.Одно из возможных предупреждений состоит в том, что общее высвобождение у животных в ответ на стрессоры может не быть критической переменной; так как авторы изучают способность к синтезу (т.е. число активных терминалов), лучшее сравнение будет с активностью популяции нейронов DA (т.е. числом активированных нейронов), которая была показана в исследованиях повторяющегося стресса Валенти и Грейс. Эти данные были бы более последовательными и менее запутанными, чем цитируемая литература, которая смешивает высвобождение, метаболиты и активацию.
Спасибо за это предложение. Мы отделили животное от человеческого труда. Мы удалили исследования, изучающие вызванное стрессом высвобождение дофамина при работе с животными, и добавили следующий текст в абзац, посвященный работе с животными:
«Исследования на животных показали, что острые стрессовые факторы, включая аверсивные стимулы, вызывают выраженную активацию дофаминовой системы с точки зрения активности популяции дофаминовых нейронов (т. Е. Количества активируемых нейронов) и в отношении поведения, индуцированного амфетамином (Valenti et al.). »
2) Какие области DA-системы активируются по сравнению с притупленными при остром или хроническом стрессе, является важной переменной. Острый и повторяющийся стресс активирует всю DA-систему, проецирующуюся на большую часть полосатого тела (Валенти и Грейс), в частности ассоциативное полосатое тело, где важна выраженность объекта, тогда как при хронической депрессии, вызванной стрессом (Холли и Мицек), притупление происходит в основном нейроны, проецирующиеся в вентромедиальное полосатое тело (Морейнс и Грейс), где обрабатываются переменные, связанные с вознаграждением.Следовательно, это разные системы, опосредующие стрессовую реакцию DA, которая варьируется в зависимости от продолжительности стрессового воздействия и индукции тревоги (острый или повторяющийся стресс) по сравнению с депрессией (хронический стресс). Это согласуется с моделью процесса оппонента Куба, где острый стресс активирует систему DA, которая при хроническом воздействии приводит к компенсаторному понижению регуляции.
Большое спасибо за то, что подняли эти важные вопросы. Мы добавили в Обсуждение следующий текст:
«Имеются данные о региональной специфичности в отношении эффектов острого и острого.хроническое стрессовое воздействие. […] Наши результаты согласуются с моделью процесса оппонента Куба, в которой острый стресс активирует дофаминовую систему, которая при хроническом воздействии приводит к компенсаторному подавлению регуляции (Koob et al., 1997) ».
3) Стресс в юношеском / подростковом возрасте имеет совсем другие эффекты, чем хронический стресс у взрослых. Следовательно, нельзя сравнивать повторяющийся стресс у взрослых с факторами стресса у подростков.
Мы отредактировали соответствующий раздел Обсуждения, чтобы прояснить, когда мы говорим о развитии и развитии.подверженность взрослому стрессу. Мы отредактировали текст следующим образом:
«Наши результаты расширяют эти выводы и согласуются с данными анима / моделей, согласно которым субкортикальная передача дофамина притупляется в ответ на несколько стрессоров во взрослом возрасте (Chrapusta, Wyatt and Masserano, 1997; Gresch et al., 1994) … Во время стресса Воздействие на животных в юношеский период и в подростковом возрасте имеет совершенно иной эффект от хронического стресса во взрослом возрасте, наши результаты также в целом согласуются с моделями стресса, обусловленного развитием (Brake et al., 2004). «
4) Обсуждение интерпретации результатов — Холли и Мицек не записывали данные с нейронов DA.
Ссылка на обзор Холли и Мичек заменена ссылкой на исследование Saal et al., 2003.
5). Раздел о материнской депривации встречается в очень раннем юношеском периоде (то есть на 7-8-е сутки послеродового периода), что намного раньше, чем ювенильный / подростковый стресс, упомянутый в статье.
Мы четко обозначили время проведения исследований материнской депривации в тексте:
«Аналогичным образом, модели ранней материнской депривации в самом раннем ювенильном периоде (начиная с 5-го дня послеродового периода) были связаны с гиподопаминергическим поведением…»
Рецензент № 2:
Авторы представляют данные о способности к синтезу дофамина (с использованием ПЭТ) у лиц с высокой подверженностью неблагоприятным воздействиям по сравнению с подобранной контрольной группой с низким уровнем неблагоприятных исходов (каждая группа с N = 17).ПЭТ-сканирование выполняется через два часа после задания на острый психосоциальный стресс (MIST), когда участники с высокой подверженностью неблагоприятным воздействиям демонстрируют повышенный опыт угрозы, но более низкое артериальное давление и более низкий уровень кортизола в ответ на тенденцию.
Меня заинтриговали эти открытия, поскольку автор связывает базальную функцию нейротрансмиттера с кумулятивными неблагоприятными воздействиями в разных случаях в неклинической популяции.
Мое волнение, однако, гаснет из-за порядка сканирования MIST и ПЭТ: если авторы ожидают воздействия стрессовых событий (хронического) на функцию дофамина, сканирование ПЭТ также будет зависеть (резко) от предшествующего стрессового опыта. .Авторам необходимо уточнить этот эффект. Поскольку дизайн не может быть изменен, как минимум было бы четкое заявление в аннотации и основном тексте.
Дизайн исследования был одинаковым для обеих групп, т.е. обе группы прошли ПЭТ-сканирование в один день с острым психосоциальным стрессом. Хотя мы не можем исключить возможность взаимодействия (см. Ниже), маловероятно, что резкие изменения в синтезе дофамина влияют на наши результаты.
В аннотацию внесены следующие изменения:
«ПЭТ-сканирование проводилось через два часа после индукции острого психосоциального стресса с использованием стресс-теста Montréal Imaging Stress Task, чтобы вызвать острый психосоциальный стресс.«
Мы также добавили следующий текст в раздел ограничений Обсуждения:
«ПЭТ-сканирование было проведено после воздействия острого психосоциального стресса. Хотя это увеличивает выброс дофамина (Pruessner et al., 2004), неясно, оказывает ли это острое влияние на способность синтеза дофамина в полосатом теле. […] необходимо исследование, чтобы определить, влияет ли острый стресс на способность к синтезу дофамина ».
Авторам необходимо предоставить анализ мощности.В этом исследовании участвовало небольшое количество субъектов (я знаю, что это может быть обычным явлением для исследований ПЭТ), поэтому авторам необходимо убедительно показать, что эффекты хорошо развиты.
Мы добавили следующий расчет мощности:
«В исследовании надежности повторного тестирования [ 18 F] -DOPA PET (Egerton et al., 2010) полосатое тело Kp er имело коэффициент внутриклассовой корреляции приблизительно 0,9 [среднее (SD) = 0,01417 (0,00127) ) мин-1 (тест) и 0,01381 (0,00127) мин-1 (повторный тест)].[…] Следовательно, для достижения мощности 0,8 с размером эффекта 1,0, a = 0,05, с использованием независимых t-критериев, потребуется n = 17 участников на группу. «
Дополнительным фактором, который необходимо учитывать (например, путем ковариации), является количество курящих в обеих группах. Несмотря на то, что количество курильщиков одинаково в обеих группах, количество курящих различается и, как было показано, влияет на нейротрансмиссию дофамина.
В текст вставлено следующее:
«Поскольку количество курения в группах было разным, и тяжелое курение может влиять на функцию дофамина (Bloomfield et al., 2014; Salokangas et al., 2000) мы выполнили ANCOVA, чтобы проверить, влияет ли курение на наши выводы. При совместном изменении количества сигарет, которые вы курили в настоящее время, разница между группами оставалась значимой в лимбическом полосатом теле только 5,2, p = 0,029, «Ép = 0,15».
В медиативном анализе было выбрано лимбическое подразделение полосатого тела (подраздел «Взаимосвязь между физиологическими и субъективными показателями»). Мотивация для этой подобласти в тексте («подключение к системе угроз») и в Материалах и методах («Исследовательские анализы проводились в функциональных подразделениях полосатого тела.») не очень сильна. Я бы посоветовал провести регрессионный анализ, чтобы определить, какой субрегион на самом деле является лучшим предсказателем для физиологических показателей.
Лимбическое полосатое тело было выбрано из-за его роли в реакции на стресс, его функциональной связи с системой обнаружения угроз и наибольших размеров эффектов для групповых различий, наблюдаемых в нашем исследовании. Однако регрессионный анализ не выявил, какой субрегион был лучшим предиктором физиологических показателей (p>.08). Поэтому мы удалили анализ посредничества из рукописи и вставили в текст следующее:
«Регрессионный анализ не выявил, какое полосатое тело / субрегион было лучшим предиктором физиологических показателей (p> 0,08)».
Рецензент № 3:
[…] Любое исследование ПЭТ требует серьезных затрат времени и ресурсов, и поэтому следует поблагодарить авторов за то, что они предприняли эти усилия. Вопрос исследования не является полностью новым, поскольку в предыдущих исследованиях изучалась возможная роль стресса в функции дофамина, в том числе с использованием ПЭТ (и авторы ссылаются на это).
Что необычно, так это сочетание факторов стресса в раннем возрасте и взрослого, чтобы создать группу с высоким совокупным пожизненным неблагоприятным воздействием; Раньше с этим не сталкивался. Можно, конечно, поспорить, представляет ли этот подход полезный подход, поскольку он смешивает эффекты ранней жизни (вероятное программирование) с более поздними острыми стрессорами, которые возникают после критических периодов развития. Фактически, некоторые авторы (например, Эллис и его коллеги с их работой по теории истории жизни) утверждают, что невзгоды ранней жизни изменяют системы стресса, чтобы лучше подготовить вас к взрослой жизни, которая, вероятно, будет похожей.В этом смысле стресс в раннем возрасте, за которым следует стресс у взрослых, может, таким образом, представлять ожидаемый результат для организма, к которому он теперь лучше подготовлен.
Как бы то ни было, я просто хотел бы более сильного обоснования конкретного выбранного экспериментального подхода, поскольку мне было неясно, что именно заставило авторов выбрать эти критерии. Кроме того, мне не сразу было ясно, почему статус этнического меньшинства считался фактором стресса для взрослых как таковым, что приравнивалось к тяжелой утрате или безработице, и я хотел бы видеть больше ссылок, оправдывающих этот подход.
Мы благодарны рецензенту за то, что он поднял эти важные моменты. Результаты эпидемиологических исследований показали, что неблагоприятные факторы раннего развития повышают риск психопатологии в ответ на стрессоры в зрелом возрасте (например, McLaughlin et al., 2010), не согласуются с теорией, согласно которой травма в детстве защищает от риска психического заболевания в ответ на взрослые стрессоры.
Мы пересмотрели обоснование нашего экспериментального подхода, включая обоснование включения статуса этнического меньшинства во Вступление, следующим образом:
«Исследования психосоциальных стрессоров и функции дофамина обычно изучали факторы риска изолированно, несмотря на то, что факторы риска группируются вместе и могут иметь общие лежащие в основе механизмы (Hjern, Wicks и Dalman; Morgan and Fisher; Wicks et al., 2005). […] Принимая во внимание данные о дофаминергической дисфункции, связанной с жестоким обращением в детстве, представленными выше, мы предположили, что здоровые люди с высоким совокупным воздействием психосоциальных стрессоров изменили бы синтез дофамина в полосатом теле по сравнению с людьми с низким воздействием ».
Мы добавили в Обсуждение:
«Мы решили набрать участников с высоким уровнем стресса в раннем развитии и в зрелом возрасте, потому что стрессоры на раннем этапе развития увеличивают риск психопатологии вслед за взрослыми стрессорами (McLaughlin et al., 2010). Однако потенциальное ограничение нашей комбинации факторов стресса в раннем развитии и во взрослом возрасте состоит в том, что оно может смешивать эффекты раннего периода жизни (то есть вероятное программирование) с острыми стрессорами в более позднем возрасте, которые возникают после периодов, чувствительных к развитию. «
Что могло быть потенциально более серьезной проблемой, было связано с методологическим подходом, выбранным авторами — если я правильно понял раздел «Материалы и методы», авторы выполнили стрессовую задачу (MIST) за два часа до сканирования ПЭТ.Предполагается ли, что стресс повлияет на синтез дофамина через два часа после его прекращения? Если бы это было так, то это должно быть предельно ясно сказано читателю, и тогда я хотел бы увидеть в рукописи раздел, где это систематически объясняется; прямо сейчас читатель остается в неведении относительно выбора именно этого обоснования. Кроме того, сравнения стресс-контроль не проводилось; испытуемые выполнили первый блок контроля, а затем непосредственно экспериментальное состояние MIST; как авторы могут быть уверены, что то, что они затем наблюдают в ПЭТ, является специфическим для стрессовой части экспериментальной манипуляции? Мне это совершенно непонятно, и мне (и я предполагаю, что и среднему читателю) было бы полезно более подробное объяснение этой части исследования.
Приносим извинения, что это было недостаточно ясно. Обоснование проведения ПЭТ-сканирования в тот же день, что и стресс-задача, состояло в том, чтобы уменьшить разницу во времени между измерениями. У нас не было предположений, что острое воздействие стресса может изменить последующую способность к синтезу дофамина. Хотя мы не можем исключить возможность того, что такие эффекты могут способствовать нашим результатам, мы не знаем доказательств того, что острый стрессор может изменить два параметра, которые влияют на наш индекс (т.е. количество доступных везикул дофамина и / или активность декарбоксилазы ароматической кислоты) в этот период времени. Обоснование для изучения взаимосвязи между показателями стресса и способностью к синтезу дофамина неявно приводится в рукописи. Мы включили эти точки следующим образом:
Материалы и методы:
«Основанием для проведения ПЭТ-сканирования в тот же день, что и стресс-задача, было уменьшение расхождения во времени между измерениями».
Обсуждение:
«Однако нам не известны свидетельства того, что острый стрессорный фактор может изменять параметры, которые влияют на наш индекс (т. Е.е. активности декарбоксилазы ароматических кислот) в сроки, использованные в нашем исследовании. «
[Примечание редакции: до принятия были запрошены дополнительные исправления, как описано ниже.]
Рукопись была улучшена, но остается одна проблема, которую необходимо решить до принятия:
Данные не позволяют утверждать, что синтез DA управляется только ранними невзгодами, потому что это может быть взаимодействие детских невзгод и манипуляции острым стрессом.Рецензенты и редактор считают, что это не снижает важности статьи, но необходимо четко указать в статье, включая заголовок (см. Ниже).
Мы благодарны профессору Бухелю и его коллегам за их дальнейший обзор. Мы внесли изменения в статью, включая название.
Рецензент № 2:
Автор ответил на мои комментарии и постарался прояснить критические моменты в рукописи.
Авторы согласны с тем, что текущее исследование не может разделить хронический и острый стресс, поэтому я проконсультируюсь с другими рецензентами, если термин «хронический» следует удалить / изменить из названия.
Благодарим рецензента. Мы убрали из названия термин «хронический». Название теперь гласит: «Влияние психосоциального стресса на дофаминергическую функцию и реакцию на острый стресс».
Рецензент № 3:
[…] По основным моментам первый вопрос, поднятый этим рецензентом, т.е. вопрос о том, почему была создана группа с высоким совокупным пожизненным неблагоприятным воздействием, получил исчерпывающий и убедительный ответ, что привело к полезному пересмотру как во Введении, так и в Обсуждении .
На второй ранее поднятый вопрос, а именно, почему стресс-задача предшествует ПЭТ-сканированию на два часа, был дан ответ, заявив, что причина заключалась в уменьшении дисперсии во времени. Здесь я менее воодушевлен тем, что это достаточный ответ. Почему важно уменьшить разброс во времени? С другой стороны, помещая эти два теста так близко друг к другу, я опасаюсь, что потенциальные последствия воздействия стрессора могли повлиять на то, что наблюдалось во время ПЭТ-сканирования.Авторы поясняют, что им не известны какие-либо доказательства, указывающие на то, что острый стрессор может влиять на параметры, рассматриваемые в ПЭТ, но, опять же, контролируемое исследование, сравнивающее стресс и отсутствие стресса до ПЭТ для изучения эффектов, вероятно, действительно влияет. не существует на данный момент, поэтому очевидно, что это не известно. Через два часа после острого стрессора может произойти любое количество вещей — например, изменения иммунной системы, геномные эффекты глюкокортикоидов, рефрактерный период чувствительности оси hpa, которые могут или не могут повлиять на последующее сканирование ПЭТ.
Таким образом, решение вызвать острый стресс так близко к ПЭТ, на мой взгляд, является проблематичным из-за отсутствия веских оснований. Просто чтобы избежать впечатления, что это не так, и чтобы другие исследователи не копировали этот дизайн, возможно, стоит добавить, что лучшим выбором было бы иметь больше времени, прошедшее после острого стрессора.
Благодарим рецензента за отзыв. Выполняя ПЭТ-сканирование так близко к стрессовой задаче, мы стремились сократить промежуток времени между заданием и ПЭТ-сканированием, чтобы уменьшить отклонения, связанные с нормальными колебаниями в способности синтеза дофамина.Мы согласны с тем, что, возможно, близость во времени задачи стресса к ПЭТ означает, что возможно, что то, что наблюдалось во время сканирования ПЭТ, могло быть затронуто задачей стресса. Хотя нам не известны какие-либо доказательства, указывающие на то, что острый стрессорный фактор может влиять на параметры, рассматриваемые в ПЭТ, для окончательного рассмотрения этой возможности необходимо контролируемое исследование, сравнивающее стресс и отсутствие стресса до ПЭТ. Учитывая это, возможно, поэтому, что оптимальный экспериментальный план позволит пройти больше времени между стрессором и ПЭТ-сканированием.Эти моменты были разъяснены в тексте ниже:
«ПЭТ-сканирование было проведено после воздействия острого психосоциального стресса. […] Тем не менее, для окончательного определения того, влияет ли острый стресс на способность к синтезу дофамина, необходимо контролируемое исследование, сравнивающее стресс и отсутствие стресса до ПЭТ. , возможно, что оптимальный экспериментальный план позволит пройти больше времени между стрессором и ПЭТ-сканированием ».
https: // doi.org / 10.7554 / eLife.46797.014
Влияние методов отвлечения внимания на боль и стресс во время родов: рандомизированное контролируемое клиническое исследование | BMC по беременности и родам
Рекомендации CONSORT были соблюдены при составлении отчетов об этом исследовании.
Дизайн исследования
Это исследование представляло собой простое слепое рандомизированное контролируемое клиническое исследование с двумя параллельными группами (интервенционная и контрольная группы). Это исследование одобрено этическим комитетом Тебризского университета медицинских наук (код: IR.TBZMED.REC.1396.453). Популяция этого исследования включает беременных женщин с гестационным возрастом 28–32 недель, которые обращались в медицинские центры города Халхал, провинция Западный Азербайджан, Иран с февраля по сентябрь 2018 года. Все участники подписали форму письменного информированного согласия. Только анализатор данных не знал о вмешательстве, полученном исследовательскими группами.
Исследуемая популяция
Критериями включения были беременность с гестационным возрастом 28–32 недели, наличие первой или второй беременности, готовность к родам в Халхальской больнице имама Хомейни и отсутствие участия в одних и тех же классах.Критериями исключения были беременность с высоким риском, включая гестационный диабет, преэклампсию, двойную и многоплодную беременность, матери с амниотической жидкостью и нарушениями плаценты, смерть плода, психические заболевания и прием определенных лекарств, наличие показаний к кесареву сечению, предыдущее кесарево сечение, аномальное предлежание плода. , стеноз таза и макросомия плода.
Рандомизация и маскирование
Участников распределили на две группы вмешательства (получатели отвлекающих методов) и контроль с помощью стратифицированной блочной рандомизации на основе количества родов (первая и вторая роды) с размерами блоков 4 и 6 и с Соотношение назначений 1: 1.Блокировка была произведена лицом, не вовлеченным в сбор и анализ данных. Чтобы скрыть выделение, тип вмешательства был написан на листе бумаги и запечатан в матовых конвертах. Конверты открывались исследователем в порядке входа участников в исследование и определялся тип группы.
Процедуры
Отбор проб начался после получения этического кодекса от этического комитета Тебризского университета медицинских наук и регистрации исследования на сайте Иранского реестра клинических испытаний.Халхал, город на северо-западе Ирана, имеет три медицинских центра. В процессе отбора проб исследователь спрашивал информацию о беременных женщинах в течение недели 28–32 через Интегрированную систему здравоохранения и связывался с матерями, у которых были некоторые критерии включения. Кроме того, по телефону исследователь кратко рассказал о плане исследования и его целях. Участники оценивались на основе критериев приемлемости, и, если они соответствовали критериям и хотели участвовать в исследовании, их просили посетить медицинский центр в определенное время.Во время посещения беременным женщинам были предоставлены исчерпывающие данные, включая цели, важность и преимущества участия в исследовании, а также этапы проведения исследования. Более того, при желании основные анкеты, включая анкету по социально-демографическим характеристикам, анкету PSS и анкету о страхе родов (W-DEQ версия A), были заполнены посредством интервью, и участники были разделены на две группы.
Консультации, основанные на методах отвлечения внимания для контроля стресса, страха и боли, были представлены участникам группы вмешательства на четырех занятиях в течение недели.Все консультации проводил первый автор. Первый сеанс проведен на 32 неделе беременности. В ходе этой встречи все участники пытались установить дружеские отношения и завоевать доверие участников. Затем исследователь подробно описал определение техники отвлечения внимания и ее влияние на обезболивание. На втором занятии группе вмешательства было объяснено несколько методов отвлечения внимания; например, просмотр фильмов, решение таблиц и головоломок, прослушивание музыки, иллюстрации будущего ребенка, запоминание памяти, обсуждение их навыков, обратный подсчет чисел, подсчет капель сыворотки, использованных во время родов, а также о личных интересах и опыте.На третьем сеансе исследователь получил отзывы о предыдущем сеансе, упражнения выполнялись дома, и участников просили выполнять упражнения, включая подсчет определенных букв во время просмотра видео и проигрывание музыки, обратный счет чисел 3 на 3 из 1000. , запишите, сколько времени они могут развлечься этими методами, и запишите свои интересы. Основываясь на их записях, исследователь предоставил им необходимое оборудование для использования во время родов. На четвертой сессии обсуждались этапы родов, ход родов, контроль стресса и страха с помощью отвлекающих приемов, пространство для родов и подготовка к родам, а также была получена обратная связь от группы вмешательства.Всех участников интервенционной и контрольной групп попросили посетить медицинский центр на 36 неделе, и W-DEQ был заполнен путем опроса их. Для участников контрольной группы после завершения W-DEQ был проведен тренинг по признакам родов, этапам родов и подходящему времени для направления в больницу. На всех занятиях неукоснительно соблюдались принципы консультирования. Затем всем участникам дали номер телефона исследователя, по которому можно связаться с исследователем в случае родовой боли и направления в больницу.Исследователь посетил больницу и использовал шкалу ВАШ для записи боли участников каждый час во время активной фазы родов. Затем анкета предполагаемого стресса была заполнена после поступления участника в родильное отделение в начале активной фазы родов посредством собеседования. Группе вмешательства, исходя из их интереса к третьему сеансу, были предоставлены отвлекающие средства, такие как фильмы, музыка, стол, головоломка, книга и многое другое. Исследователь был активен вместе с матерью в течение всей фазы активной фазы и всех техник отвлечения внимания участников, включая обратный подсчет чисел, подсчет капель сыворотки, запоминание, иллюстрирование и т. Д.проводились в присутствии исследователя. Матерям показали их любимый фильм, и их попросили внимательно посмотреть фильм и сосчитать определенные буквы при просмотре. Контрольная группа получала плановую помощь.
Первичные исходы
Первичные исходы включали выраженность родовой боли и воспринимаемый стресс, которые измерялись с помощью ВАШ и PSS, соответственно, до вмешательства и в активной фазе родов. ВАШ — это шкала длиной 10 см, с помощью которой пациент должен определить свою собственную оценку боли по этой шкале от нуля (безболезненно) до 10 (самая сильная боль, которую можно представить).По этой шкале нулевой балл означает роды без боли, 1–3 — легкие, 6–4 — средние, 9–7 — сильные и 9–10 — очень сильные боли [30]. PSS состоит из 14 пунктов, а оценки основаны на 5 пунктах Лайкерта следующим образом: никогда = 0, почти никогда = 1, иногда = 2, часто = 3 и много раз = 4 балла. Пункты 4–5–6-7, 9, 10 и 13 оцениваются в обратном порядке (никогда = 4, много раз = 0). Наименьший балл — 0, наивысший балл — 56. Более высокий балл указывает на более выраженный стресс [31]. Надежность персидской версии этого вопросника была рассчитана Bastani et al., методом внутренней согласованности. Для этого вопросника они получили коэффициент альфа Кронбаха 74% [32].
Вторичные исходы
Вторичные исходы включали боязнь родов, продолжительность активной фазы родов и второго периода родов, общую продолжительность родов, оценку по шкале Апгар на первой минуте и потребление окситоцина. W-DEQ-Version A использовался для оценки страха перед родами перед вмешательством и снова на 36-й неделе беременности. В анкете 33 вопроса.Матери определяют свои личные чувства на основе 6-балльной шкалы Лайкерта (вообще = 0, очень низкий = 1, низкий = 2, средний = 3, высокий = 4 и очень высокий = 5). Вопросы 1, 4, 5, 9, 10, 13, 14–16–17–18, 21–22–23, 26, 28–29–30 оцениваются в обратном порядке. Диапазон баллов составляет от 0 до 165, и более высокий балл указывает на больший страх [33]. Надежность персидской версии этого вопросника была оценена Абеди и др., Которые сообщили, что альфа Кронбаха составляет 0,64 [34].
Продолжительность родов, оценка по шкале Апгар на первой минуте и потребление окситоцина были записаны исследователем в партограмме во время схваток и родов.
Статистический анализ
Размер выборки в этом исследовании был рассчитан на основе переменных боли и стресса с использованием программного обеспечения G-Power. По результатам исследования Madadi et al. (2016) относительно переменной боли, принимая m 1 = 8,9 (боль до вмешательства), m 2 = 7,9 (боль после вмешательства), sd 1 = 1,2, sd 2 = 0,9, α = 0,05 и Power = 95% были рассчитаны как 31 [35]. По результатам Mirghafourvand et al.(2014) по переменной воспринимаемого стресса и с учетом m 1 = 26,2 (воспринимаемый стресс до вмешательства), со снижением на 20% средней оценки воспринимаемого стресса в результате вмешательства (m 2 = 19,65), sd 1 = Sd 2 = 5,5, α = 0,05 и мощность = 95% было рассчитано как 28 [36]. Поскольку размер выборки был рассчитан на основе переменной боли, был больше, с учетом 10% отсева, окончательный размер выборки был рассчитан как 34.
Статистический анализ настоящего исследования был проведен с использованием программного обеспечения SPSS 24.Нормальность количественных данных исследовали с помощью критерия Кормогрова-Смирнова. Результаты показали, что продолжительность второго периода родов и оценка боли не имели нормального распределения . Критерии хи-квадрат, хи-квадрат для тренда, независимые t и точные тесты Фишера использовались для оценки согласованности этих двух групп с точки зрения социально-демографических характеристик. Для сравнения средней оценки воспринимаемого стресса и страха при родах использовался независимый t-тест до вмешательства и тест ANCOVA с корректировкой исходных значений и фактор стратификации (первые или вторые роды) после вмешательства.Для сравнения продолжительности активной фазы и общей продолжительности родов использовался независимый t-критерий, а для сравнения продолжительности второй стадии родов и средней боли во время родов в двух группах использовался U-критерий Манна-Уитни. . Тест хи-квадрат применялся для сравнения частоты потребления окситоцина в двух группах вмешательства и контрольной группе.