
Содержание
как определить парный и непарный , мягкий и твердый, глухой и звонкий согласный звук
математика пятый класс первая часть
математика пятый класс первая часть
35 баллов!
Найдите сложное предложение с сочинительной и подчинительной связью между частями.
Выпишите его (5 баллов).
Подчеркните основы, выделите со
…
юзы / союзные слова, соединяющие эти части (5 баллов).
Что неясно представляешь, то неясно и выскажешь: неточность и запутанность выражений свидетельствуют о запутанности мыслей.
В тяжёлые дни войны или каких-нибудь бедствий жители средневековых городов собирались в главном соборе, чтобы обсудить важные дела, там же хранились запасы оружия и продовольствия.
Известно, что древние греки, у которых было горе, приходили к статуе Зевса, созданной скульптором Фидием, и находили утешение, глядя на лицо бога — таким оно было добрым и прекрасным.
Законы перспективы не изменились и сейчас, спустя почти пять веков со времени их открытия, и все художники неизменно пользуются ими, когда пишут картины или рисуют.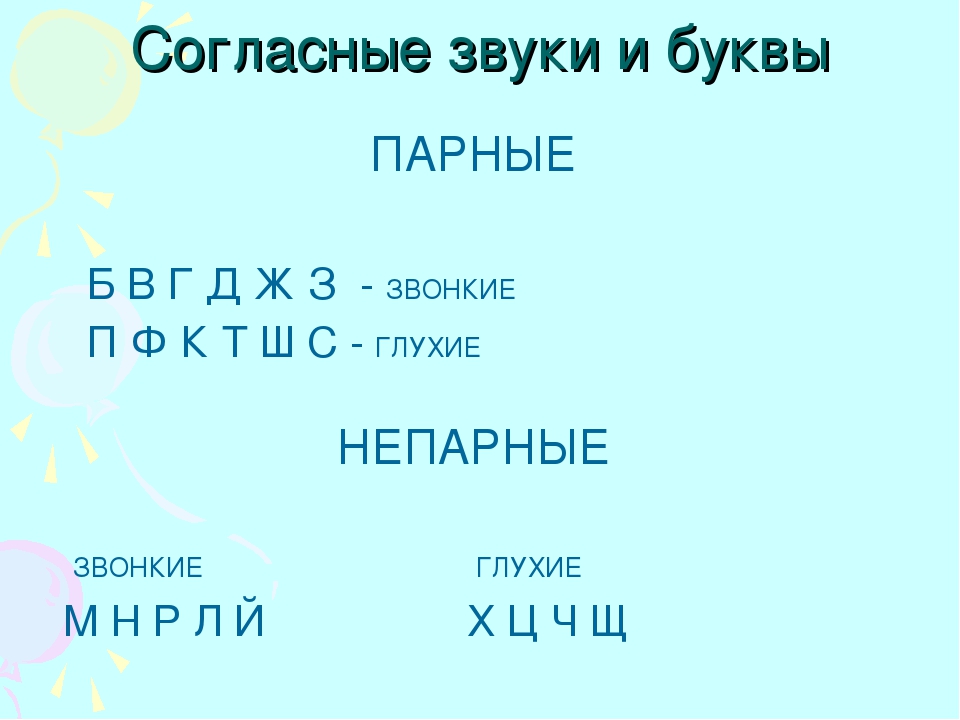
Обычно с ним гуляла хозяйская дочь Тамара.
Предлог с(?) употреблён в творительном
падеже(?) выражает
отношения (?).
1) идти вдоль дороги; 2) ни рыба ни мясо; 3) да здравствует солнце лучистое; 4) спрятались за домом; 5) видит да слышит
Какие конструкции с союзом? С
…
Предлогом? С частицей?
Все знают о писателе Александре Грине все восхищаются этим романтиком с пылкой творческой фантазией. РАССТАВИТЬ ЗНАКИ ПРЕПИНАНИЯ И СИНТАКТИЧЕСКИЙ РАЗБ
…
ОР!!! БЫСТРО!!!! ПОЖАЛУЙСТА
«Как красиво» , как — это союз?
Срочно !!!!!!!Найдите предложения, которые соединяются с предыдущими с помощью ука-занного средства: 1) тематическая группа слов; 2) повторы слов и уп
…
отребление однокоренных слов; 3) синонимы; 4) антонимы. Напишите номера этих предло-жений.1. Василий Кириллович Авраменко родился в 1895 г. на Черкас-чине и известен среди украинской диаспоры в Америке как «отец украинского танца». 2. Прибыв в Торонто 2 декабря 1925, танцовщик организует по городам Канады и Америки школы украинских танцев.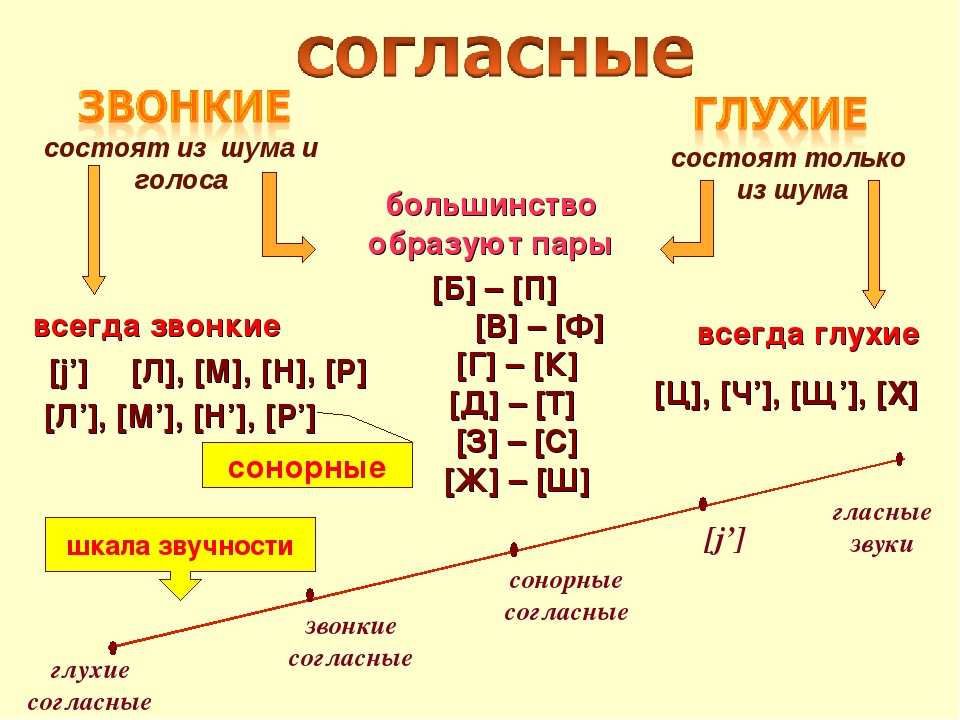 3. Под заголовком «Украинский балет» в 1936 г. дал первый концерт с учениками, который прошел с небывалым успехом. 4. 25 мая 1931 г. остоялось большое выступление украинских артистов на сцене Метро-политен-опера в Нью-Йорке, в котором украинцы показали величие и красоту творческой души своего народа. 5. На сцену В. К. Аврамен-ко вывел 600 танцоров, 100 хористов и оркестр. 6. Успех был потря-сающий. В прессе не умолкали похвалы в адрес артистов-украинцев. 7. Умер В. Авраменко в Нью-Йорке 6 мая 1981 года. 8. В своем за-вещании он просил перевести его прах в Украину, в родной Стеблев на Черкасчине. 9. Выполняя волю артиста, его прах в 1993 г. был пе-ревезен на родину. 10. В. К. Авраменко вывел украинский танец на мировую арену.
3. Под заголовком «Украинский балет» в 1936 г. дал первый концерт с учениками, который прошел с небывалым успехом. 4. 25 мая 1931 г. остоялось большое выступление украинских артистов на сцене Метро-политен-опера в Нью-Йорке, в котором украинцы показали величие и красоту творческой души своего народа. 5. На сцену В. К. Аврамен-ко вывел 600 танцоров, 100 хористов и оркестр. 6. Успех был потря-сающий. В прессе не умолкали похвалы в адрес артистов-украинцев. 7. Умер В. Авраменко в Нью-Йорке 6 мая 1981 года. 8. В своем за-вещании он просил перевести его прах в Украину, в родной Стеблев на Черкасчине. 9. Выполняя волю артиста, его прах в 1993 г. был пе-ревезен на родину. 10. В. К. Авраменко вывел украинский танец на мировую арену.
Списать предложения, вставить пропущенные буквы. Указать спряжение глаголов, подчеркнуть глаголы-исключения.Заунывный ветер гон_т стаю туч на край неб
…
ес. (Н. Некрасов.) Ветер по морю гуля_т и кораблик подгоня_т. (А. Пушкин.) Легко дыш_т грудь. Тывдыха_шь запах полевых цветов. С высоты зорко смотр_т на землю орёл. Он высматрива_т добычу.пожалуйста помогите
С высоты зорко смотр_т на землю орёл. Он высматрива_т добычу.пожалуйста помогите
звуко буквенный разбор слова площадь плиз Help meee
Как обозначается парный глухой. Парные звонкие и глухие согласные. Парные и непарные глухие согласные
При произнесении звонких
согласных поток воздуха создает колебания голосовых связок.
Если голосовые связки не задействованы, то звук считается глухим
.
Но в русском языке звонкая буква не всегда обозначает звонкий звук (и наоборот: глухой согласный не всегда обозначает глухой звук). Это зависит от положения буквы в слове.
Звонкий согласный часто оглушается в конце слова
. Например в слове «дуршлаг» мы на конце читаем «к», потому что звук в слабой позиции. Также может оглушаться перед глухим согласным
. Например, слове «походка» мы произносим как «пахот
ка».
Чтобы определить, какую букву будет правильно написать, букву нужно поставить в однокоренном слове в сильную позицию (то есть перед гласной или согласными М, Л, Н, Р).
Например: «лодка» — «лодочка», «гриб» — «грибной».
Таблица
Парные | Звонкие | Глухие |
| Б | ||
| Ф | ||
| Г | ||
| Т | ||
| Ж | ||
| С | ||
Непарные | Л, М, Н, Р, Й (сонорные) |
Также парными по глухости/звонкости являются пары мягких согласных, от указанных в таблице. Например: «б’ — п’», «в’ — ф’».
Твердые и мягкие
В словах одна и та же буква может обозначать и твердые и мягкие звуки.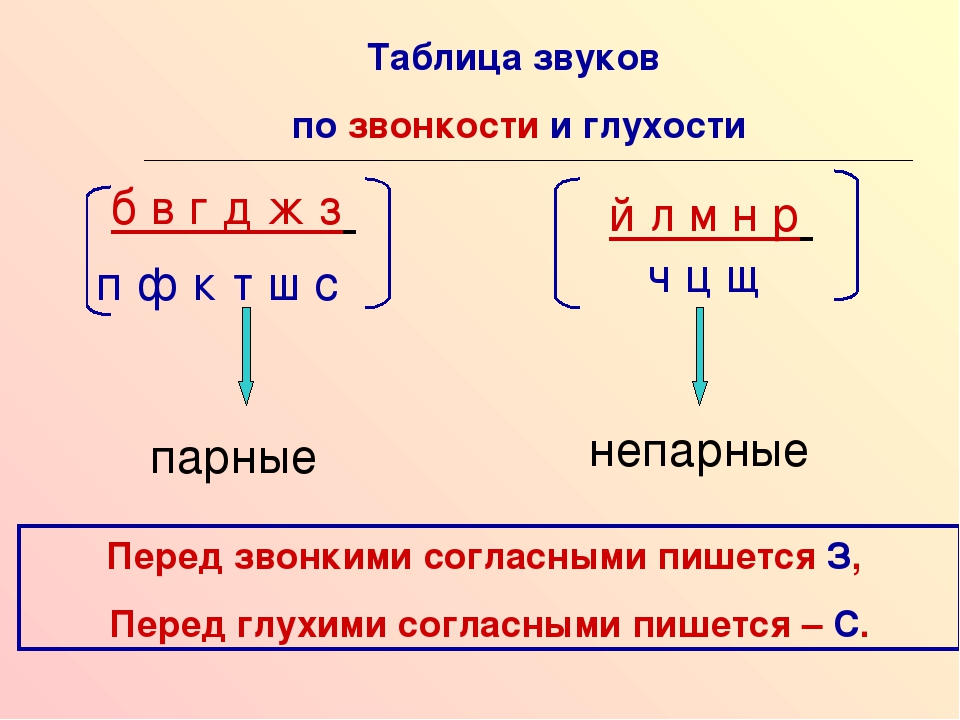 Это связано с влиянием последующих согласных на мягкость/твердость. Перед А, О, У, Ы, Э звучат твердые согласные, перед И, Е, Ё, Ю, Я — мягкие.
Это связано с влиянием последующих согласных на мягкость/твердость. Перед А, О, У, Ы, Э звучат твердые согласные, перед И, Е, Ё, Ю, Я — мягкие.
Таблица
Парные Перед А, О, У, Ы, Э – твердые. Перед И, Е, Ё, Ю, Я – мягкие. | Твердые | Мягкие | ||
| б | б ‘ | белый | ||
| ваза | в | в ‘ | ||
г | ||||
| д | д ‘ | дядя | ||
| зола | з | з ‘ | ||
| к | к ‘ | кирпич | ||
| лак | л | л ‘ | ||
| м | м ‘ | мир | ||
| наш | н | н ‘ | ||
| п | п ‘ | песня | ||
| роза | р | р ‘ | ||
| с | с ‘ | синий | ||
| туча | т | т ‘ | ||
| ф | ф ‘ | фильм | ||
| холка | х | х ‘ | Хельсинки | |
Непарные | ж, ш, ц | ч, щ, й | ||
На этом уроке мы научимся различать звонкие и глухие согласные звуки и обозначать их на письме согласными буквами.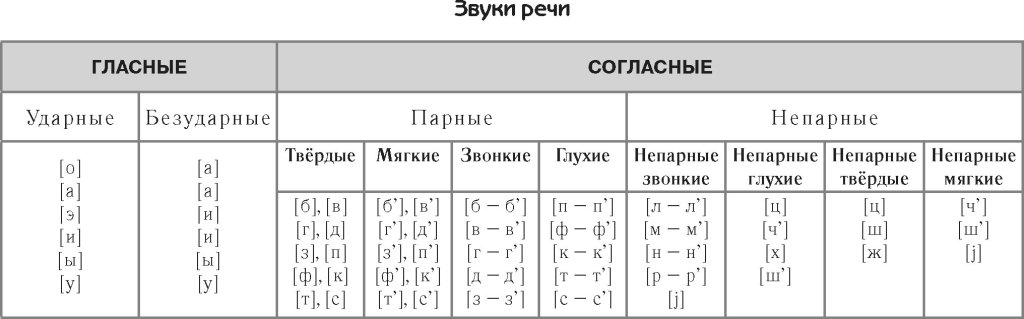 Узнаем, какие согласные называют парными и непарными по звонкости — глухости, сонорными и шипящими.
Узнаем, какие согласные называют парными и непарными по звонкости — глухости, сонорными и шипящими.
Звонкие и глухие согласные звуки
Вспомним, как рождаются звуки речи. Когда человек начинает говорить, то выдыхает воздух из лёгких. По дыхательному горлу он бежит в узенькую гортань, где находятся особые мышцы — голосовые связки. Если человек произносит согласные звуки, то закрывает (хоть немного) рот, из-за этого получается шум. Но шумят согласные по-разному.
Проведем опыт: зажмем уши и произнесем звук [п], а потом звук [б]. Когда мы произнесли звук [б], связки натянулись и начали дрожать. Это дрожание превратилось в голос. В ушах немного зазвенело.
Можно провести похожий опыт, положив руки на шею с правой и левой стороны, и произнести звуки [д] и [т]. Звук [д] произносится намного звонче, более звучно. Такие звуки учёные назвали звонкими
, а звуки, которые состоят только из шума — глухими.
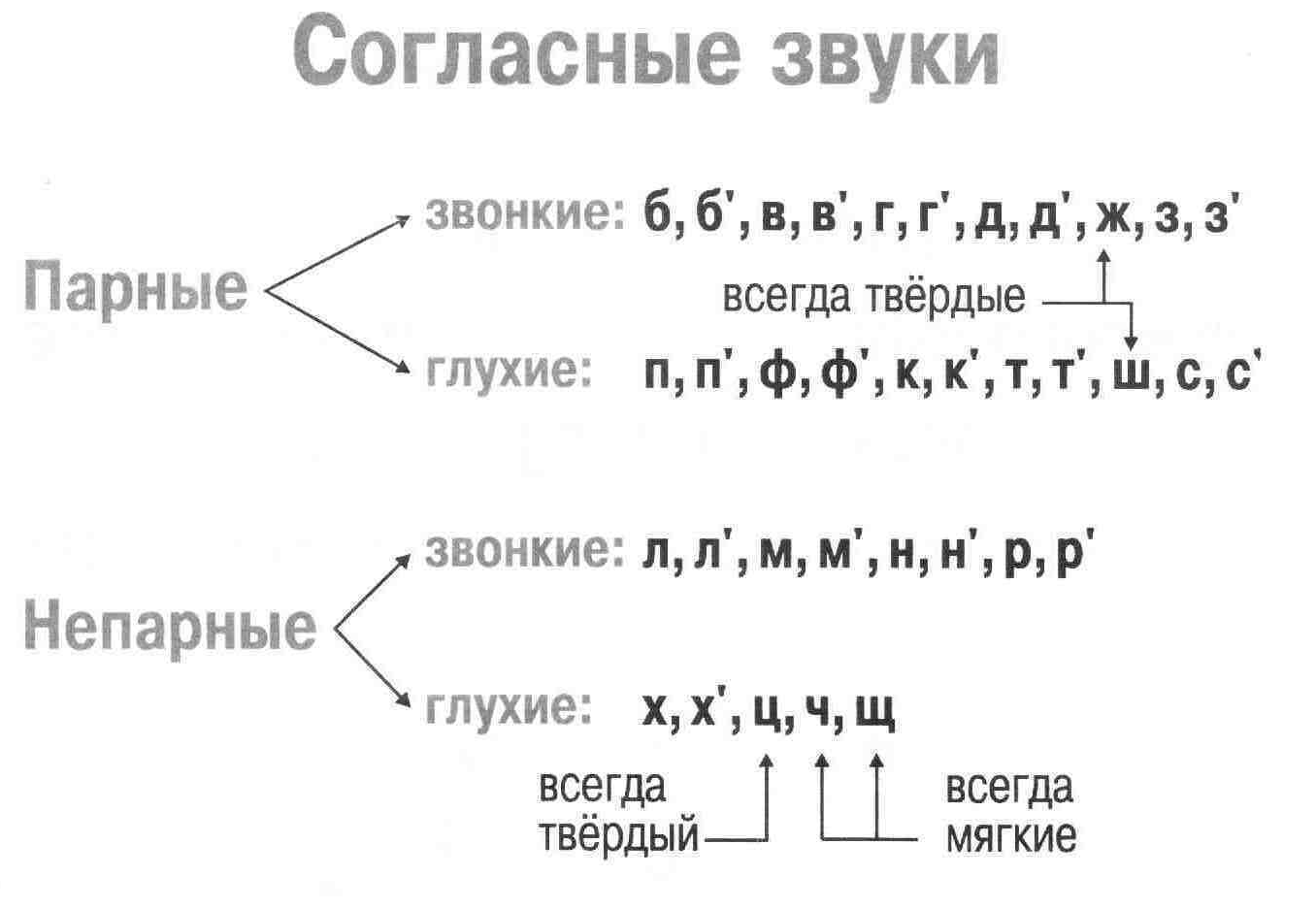
Парные по звонкости-глухости согласные звуки
Попробуем разделить звуки на две группы по способу произношения. Заселим фонетические домики в городе звуков. Договоримся: на первом этаже будут жить глухие звуки, а на втором — звонкие. Жители первого домика:
Заселим фонетические домики в городе звуков. Договоримся: на первом этаже будут жить глухие звуки, а на втором — звонкие. Жители первого домика:
Эти согласные звуки называются парными
по звонкости — глухости.
Рис. 1. Парные звонкие и глухие согласные звуки ()
Они очень похожи друг на друга — настоящие «двойняшки», их и произносят почти одинаково: одинаково складываются губы, одинаково двигается язык. Но у них есть пары и по мягкости — твёрдости. Добавим их в домик.
У звуков [ж] и [ш] нет парных мягких звуков, они всегда твёрдые
. А ещё их называют шипящими
звуками.
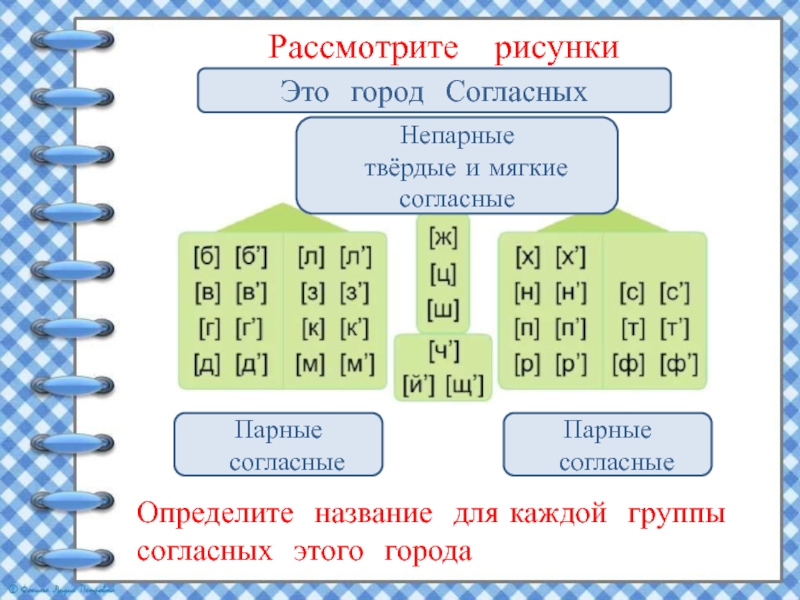
Все эти звуки обозначаются буквами:
Непарные звонкие согласные звуки
Но не все согласные звуки и буквы образуют пары. Те согласные, которые пар не имеют, называют непарные.
Поселим непарные согласные звуки в свои домики.
Во второй домик — непарные
звонкие согласные
звуки:
Напомним, что звук [й’] всегда только мягкий.
Поэтому в нашем домике он будет жить один. Эти звуки обозначают на письме буквами:
Звуки второго домика называют ещё сонорными
, потому что образуются они с помощью голоса и почти без шума, они очень звучные. Слово «сонорный» в переводе с латинского «сонорус» означает звонкий.
Непарные глухие согласные звуки
В третий домик мы поселим непарные глухие согласные
звуки:
Вспомним, что звук [ц] всегда твёрдый
, а [ч’] и [щ’] — всегда мягкие.
Непарные глухие согласные звуки обозначают на письме буквами:
Звуки [ч’], [щ’] — шипящие
звуки.
Вот мы и заселили наш город согласных звуков и букв. Теперь сразу понятно, почему согласных букв — 21, а звуков — 36.
Рис. 2. Звонкие и глухие согласные звуки ()
Закрепление знаний на практике
Выполним задания.
1. Рассмотрим картинки и превратим одно слово в другое, заменив только один звук. Подсказка: вспомним пары согласных звуков.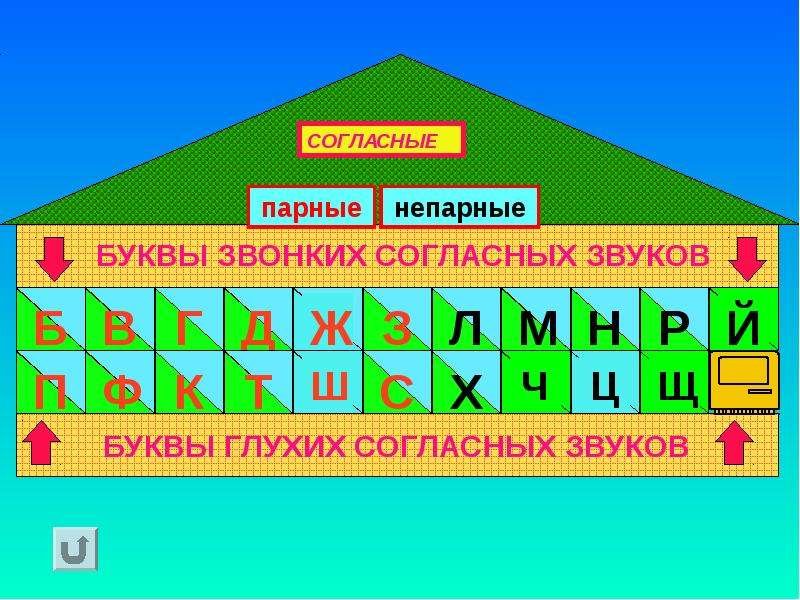
д
очка — точка
б
очка — почка
ш
ар — жар
удочка — уточка
2. Существуют загадки, смысл которых заключён в знании согласных звуков, они называются шарады. Попробуйте их отгадать:
1) С глухим согласным наливаюсь в поле,
Со звонким — сам звеню я на раздолье. (Колос — голос
)
2) С глухим — траву она срезает,
Со звонким — листья объедает. (Коса — коза)
3) С «эм» — приятен, золотист, очень сладок и душист.
С буквой «эль» зимой бывает, а весною исчезает. (Мёд- лёд)
Для того чтобы развить умение произносить какие-то звуки, особенно шипящие, учат скороговорки. Скороговорку рассказывают сначала медленно, а потом ускоряют темп. Попробуем выучить скороговорки:
- Шесть мышат в камышах шуршат.
- У ежа — ежата, у ужа — ужата.
- Два щенка щека к щеке грызли щётку в уголке.

Итак, сегодня мы узнали, что согласные звуки могут быть звонкими и глухими и как обозначаются на письме эти звуки.
- Андрианова Т.М., Илюхина В.А. Русский язык 1. М.: Астрель, 2011. ().
- Бунеев Р.Н., Бунеева Е.В., Пронина О.В. Русский язык 1. М.: Баллас. ().
- Агаркова Н.Г., Агарков Ю.А. Учебник по обучению грамоте и чтению: Азбука. Академкнига/Учебник.
- Fictionbook.ru ().
- Deafnet.ru ().
- Samouchka.com.ua ().
- Андрианова Т.М., Илюхина В.А. Русский язык 1. М.: Астрель, 2011. Стр. 38, упр. 2; Стр. 39, упр. 6; Стр. 43, упр. 4.
- Сосчитайте, сколько звонких согласных и сколько глухих в слове неудовлетворительный
?
(Звонких согласных — 9 — Н, Д, В, Л, В, Р, Л, Н, Й, различных -6, глухих согласных — 2 — Т, Т, различных — 1.)
. - Прочитайте пословицу: «
Умей вовремя сказать, вовремя смолчать».
Назовите буквы, которые обозначают звонкие согласные звуки. (Звонкие согласные звуки обозначают в пословице буквы М, Й, В, Р, З, Л.)
(Звонкие согласные звуки обозначают в пословице буквы М, Й, В, Р, З, Л.)
- 4* Используя полученные на уроке знания, напишите сказку или нарисуйте комикс на тему «В городе согласных звуков».
Как известно, звуки речи можно разделить на гласные (произносимые только голосом) и согласные (в их произнесении участвует шум). Многие согласные можно разбить на пары по их характеристикам, но не все.
Парные и непарные согласные по глухости-звонкости
Сразу надо оговориться, что таких звуков, которые непарные по всем признакам, всего четыре. Мы поговорим о них в конце статьи. Большинство же по одному признаку входит в пару, а по другому — нет. Поэтому бессмысленно писать про согласный «непарный» – надо указать, по какому признаку.
Согласные различаются по глухости-звонкости. Это означает, что при произнесении одних из них используется больше голоса (сонорные, звонкие), а в других больше шума (глухие) или даже вообще один шум (шипящие).
Сонорные — это очень звонкие согласные, голоса в них много, а шума мало.
Два сонорных согласных — [Л] и [Р] – даже могут при некоторых обстоятельствах образовывать слог, то есть вести себя как гласные. Наверняка вы встречали ошибочное написание «театор». Оно объясняется как раз тем, что [Р] в этом слове слогообразующее. Другие примеры — слова «Александр», «смысл».
Непарные звонкие согласные звуки — это как раз сонорные. Их пять:
Иногда [Й] не относят к сонорным, но он все равно остается звонким непарным. Посмотрим на таблицу.
По ней видно, что, кроме звонких непарных, имеются и звуки, являющиеся непарными глухими. Большинство из них шипящие; к шипящим не относится только глухой непарный согласный звук [Ц].
В этой статье мы рассматриваем лишь русские звуки речи. В других языках распределение по парам может быть иным. Например, в тибетском языке имеется глухая пара к звонкому [Л].
Пары по твердости-мягкости
Кроме глухости-звонкости, русские согласные образуют пары по твердости-мягкости.
Это значит, что одни из них на слух воспринимаются как более мягкие. Тогда мы обычно как-то обозначаем это на письме: например, пишем мягкий знак или одну из гласных Е, Ё, Ю, Я.
Тогда мы обычно как-то обозначаем это на письме: например, пишем мягкий знак или одну из гласных Е, Ё, Ю, Я.
Устная речь первична (любому понятно, что она появилась раньше письменной), поэтому неправильно говорить: «Звук [Н’] в слове КОНЬ мягкий, потому что после него стоит Ь». Напротив, мы пишем Ь, потому что Н’ мягкий.
По твердости-мягкости согласные также составляют пары. Но и в этом случае не все. В русском языке есть непарные мягкие и непарные твердые согласные звуки.
Непарные твердые согласные звуки — это, в основном, шипящие ([Ж], [Ш]) и [Ц]. Они всегда образуются у дальнего нёба.
А вот в предке нашего языка, старославянском, напротив, [Ж] и [Ш] были всегда мягкими и не имели твердой пары. Тогда мягкими не бывали [К], [Г] и [Х]. В настоящее время можно встретить (некогда единственно возможное) произношение с мягким [Ж’] [ДРОЖ’Ж’И] или [ДОЖ’] (дождь), но это теперь необязательно.
Непарные мягкие — это [Й’] и опять же шипящие [Ч’] и [Щ’].
То есть все шипящие являются или всегда твердыми, или всегда мягкими.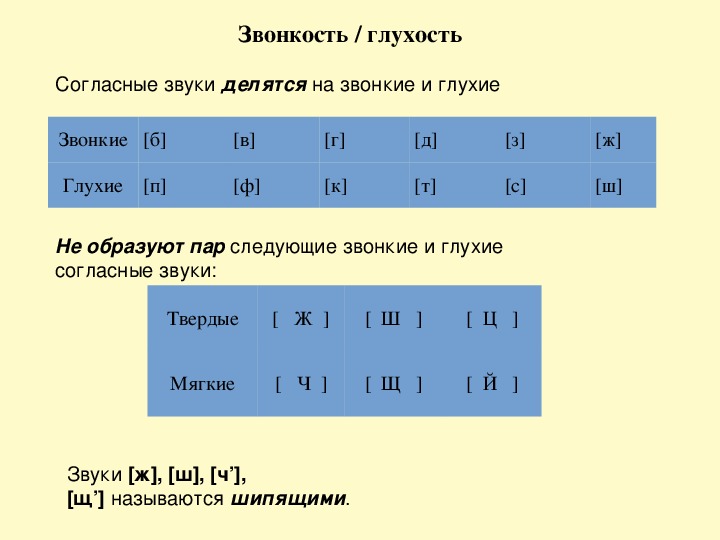 Буква Ь после них не указывает на мягкость, она выполняет грамматическую функцию (например, даже не зная, что такое «плешь», любой сразу скажет, что это слово женского рода, ведь в мужском роде после шипящих Ь не ставится). Твердые непарные шипящие согласные звуки в слове могут иметь при себе Ь, но это не значит, что они должны смягчиться. Это значит, что перед нами существительное 3 склонения, наречие или глагол.
Буква Ь после них не указывает на мягкость, она выполняет грамматическую функцию (например, даже не зная, что такое «плешь», любой сразу скажет, что это слово женского рода, ведь в мужском роде после шипящих Ь не ставится). Твердые непарные шипящие согласные звуки в слове могут иметь при себе Ь, но это не значит, что они должны смягчиться. Это значит, что перед нами существительное 3 склонения, наречие или глагол.
Непарные мягкие согласные звуки в слове вызывают желание поставить после них Ь, что часто не требуется. Поэтому есть смысл запомнить, что в сочетаниях ЧК, ЧН и т.п. Ь после Ч не нужен.
Звуки «совсем непарные»
В русском языке большинство согласных или парные по обоим признакам, или парные по одному признаку и непарные по другому. Например
, в слове [П’ЭН’] (пень) звук [П’] парный и по глухости-звонкости (П’ – Б’), и по твердости-мягкости (П’ — П), а звук [Н’] парный по твердости-мягкости (Н’ — Н), но непарный по глухости-звонкости.
Однако есть несколько звуков, непарных по обоим признакам. Это звуки [Й’] (непарный звонкий, непарный мягкий), [Ч’] (непарный мягкий, непарный глухой), [Щ’] (непарный мягкий, непарный глухой) и [Ц] (непарный твёрдый, непарный глухой). Такие звуки нередко загадывают в олимпиадах по русскому языку. Например,
Это звуки [Й’] (непарный звонкий, непарный мягкий), [Ч’] (непарный мягкий, непарный глухой), [Щ’] (непарный мягкий, непарный глухой) и [Ц] (непарный твёрдый, непарный глухой). Такие звуки нередко загадывают в олимпиадах по русскому языку. Например,
«Угадайте звук по характеристике: непарный твёрдый, непарный глухой». Мы уже видим, что это [Ц].
Что мы узнали?
Из статьи про парные и непарные согласные, мы узнали, что в русском языке существуют как парные, так и непарные согласные. Парные согласные различаются по глухости-звонкости и по твердости-мягкости.
Тест по теме
Оценка статьи
Средняя оценка: 4.1
. Всего получено оценок: 103.
Современный русский алфавит состоит из 33 букв. Фонетика современного русского числа определяет 42 звука. Звуки бывают гласные и согласные. Буквы ь (мягкий знак) и ъ (твёрдый знак) не образуют звуков.
Гласные звуки
В русском языке 10 гласных букв и 6 гласных звуков.
- Гласные буквы: а, и, е, ё, о, у, ы, э, ю, я.
- Гласные звуки: [а], [о], [у], [э], [и], [ы].
Для запоминания гласные буквы часто записывают парами по сходному звучанию: а-я, о-ё, е-э, и-ы, у-ю.
Ударные и безударные
Число слогов в слове равно числу гласных в слове: лес — 1 слог, вода — 2 слога, дорога — 3 слога и т.д. Слог, который произносится с большей интонацией, является ударным. Гласная, образующая такой слог, является ударной, остальные гласные в слове — безударными. Положение под ударением называют сильной позицией, без ударения — слабой позицией.
Йотированные гласные
Значимое место занимают йотированные гласные — буквы е, ё, ю, я, которые означают два звука: е → [й’][е], ё → [й’][о], ю → [й’][у], я → [й’][а]. Гласные являются йотированными в том случае, если:
- стоят в начале слова (ель, ёлка, юла, якорь),
- стоят после гласного (какое, поёт, заяц, каюта),
- стоят после ь или ъ (ручье, ручьём, ручью, ручья).
В остальных случаях буквы е, ё, ю, я означают один звук, но однозначного соответствия нет, так как различные позиции в слове и различные сочетания с согласными этих букв рождают разные звуки.
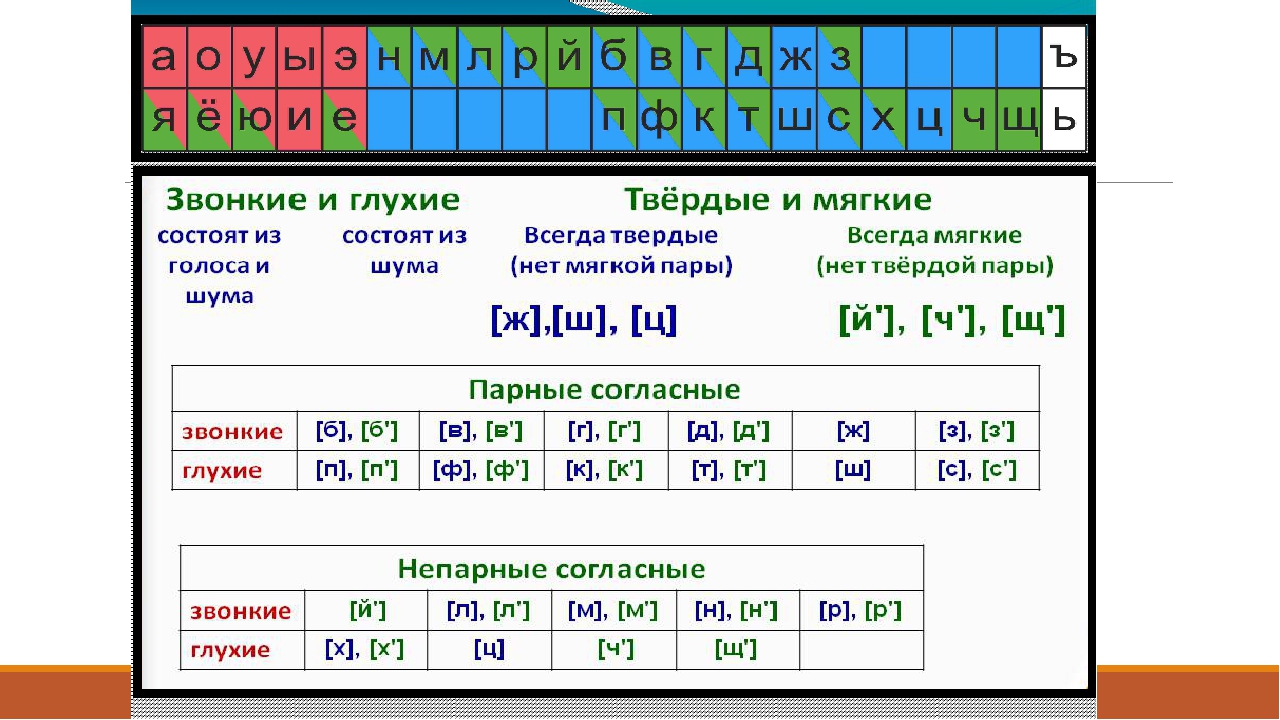
Согласные звуки
Всего 21 согласная буква и 36 согласных звуков. Несоответствие в количестве означает, что некоторые буквы могут означать разные звуки в разных словах — мягкие и твёрдые звуки.
Согласные буквы: б, в, г, д, ж, з, й, к, л, м, н, п, р, с, т, ф, х, ц, ч, ш, щ.
Согласные звуки: [б], [б’], [в], [в’], [г], [г’], [д], [д’], [ж], [з], [з’], [й’], [к], [к’], [л], [л’], [м], [м’], [н], [н’], [п], [п’], [р], [р’], [с], [с’], [т], [т’], [ф], [ф’], [х], [х’], [ц], [ч’], [ш], [щ’].
Знак ‘ означает мягкий звук, то есть буква произносится мягко. Отсутствие знака говорит о том, что звук твёрдый. Так, [б] — твёрдый, [б’] — мягкий.
Звонкие и глухие согласные
По тому, как мы произносим согласные звуки, существует разница. Звонкие согласные образуются в сочетании голоса и шума, глухие согласные образуются за счёт шума (голосовые связки не вибрируют). Всего 20 звонких согласных звуков и 16 глухих согласных звуков.
| Звонкие согласные | Глухие согласные | ||
|---|---|---|---|
| непарные | парные | парные | непарные |
| й → [й»] | б → [б], [б»] | п → [п], [п»] | ч → [ч»] |
| л → [л], [л»] | в → [в], [в»] | ф → [ф], [ф»] | щ → [щ»] |
| м → [м], [м»] | г → [г], [г»] | к → [к], [к»] | ц → [ц] |
| н → [н], [н»] | д → [д], [д»] | т → [т], [т»] | х → [х], [х»] |
| р → [р], [р»] | ж → [ж] | ш → [ш] | |
| з → [з], [з»] | с → [с], [с»] | ||
| 9 непарных | 11 парных | 11 парных | 5 непарных |
| 20 звонких звуков | 16 глухих звуков | ||
По парности-непарности звонкие и глухие согласные делятся на:
б-п, в-ф, г-к, д-т, ж-ш, з-с
— парные по звонкости-глухости.
й, л, м, н, р
— всегда звонкие (непарные).
x, ц, ч, щ
— всегда глухие (непарные).
Непарные звонкие согласные называются сонорными.
Среди согласных по уровню «шумности» также выделяют группы:
ж, ш, ч, щ
— шипящие.
б, в, г, д, ж, з, к, п, с, т, ф, х, ц, ч, ш, щ
— шумные.
Твёрдые и мягкие согласные
| Твёрдые согласные | Мягкие согласные | ||
|---|---|---|---|
| непарные | парные | парные | непарные |
| [ж] | [б] | [б»] | [ч»] |
| [ш] | [в] | [в»] | [щ»] |
| [ц] | [г] | [г»] | [й»] |
| [д] | [д»] | ||
| [з] | [з»] | ||
| [к] | [к»] | ||
| [л] | [л»] | ||
| [м] | [м»] | ||
| [н] | [н»] | ||
| [п] | [п»] | ||
| [р] | [р»] | ||
| [с] | [с»] | ||
| [т] | [т»] | ||
| [ф] | [ф»] | ||
| [х] | [х»] | ||
| 3 непарных | 15 парных | 15 парных | 3 непарных |
| 18 твёрдых звуков | 18 мягких звуков | ||
Какие звуки называют согласными?
Из чего состоит согласный звук?
Какими бывают согласные звуки?
Сколько в русском алфавите согласных букв и согласных звуков?
Какие согласные звуки всегда твёрдые, а какие — всегда мягкие?
Какими буквами обозначают мягкость согласного звука?
Звуки, при произношении которых воздух встречает во рту препятствие, называют согласными звуками
. Согласный звук состоит из шума и голоса или только из шума.
Согласный звук состоит из шума и голоса или только из шума.
Согласные звуки делятся на звонкие и глухие
. Звонкие состоят из шума и голоса, глухие — только из шума.
Только из шума состоят звуки: [к], [п], [с], [т], [ф], [х], [ц], [ч], [ш], [щ]. Это глухие согласные звуки.
Многие согласные звуки образуют пары по звонкости-
глухости
: [б]—
[п], [в]—
[ф], [г]—
[к], [д]—
[т], [з]—
[с], [ж]—
[ш].
Для запоминания звонких согласных можно выучить фразу: «У ЛьВа и ЖаБы МНоГо ДРуЗеЙ
».
См. все фразы для запоминания звонких и глухих согласных .
Глухие согласные легко запомнить по фразе: «СТёПКа, ХоЧеШь ЩеЦ?
—
Фу!
».
Согласные звуки обозначаются буквами:
Б
, В
, Г
, Д
, Ж
, З
, Й
, К
, Л
, М
, Н
, П
, Р
, С
, Т
, Ф
, Х
, Ц
, Ч
, Ш
, Щ
.
Всего в русском языке есть 21 согласная буква
.
Согласные звуки бывают также твёрдыми и мягкими.
Твёрдые и мягкие звуки
различаются положением языка при произнесении. При произнесении мягких согласных средняя спинка языка поднята к твёрдому нёбу.
Большинство согласных звуков образуют пары по твёрдости-мягкости:
Не образуют пар по твёрдости-мягкости следующие твёрдые и мягкие согласные звуки:
| Твёрдые | [ж] | [ш] | [ц] |
|---|---|---|---|
| Мягкие | [ч❜] | [щ❜] | [й❜] |
Таблица «Согласные звуки: парные и непарные, звонкие и глухие, твёрдые и мягкие» (1-4 классы)
Примечание:
в начальной школе твёрдые согласные звуки обозначаются синим цветом, мягкие согласные звуки — зелёным цветом, гласные звуки — красным цветом.
Твёрдость
согласных звуков обозначается на письме гласными буквами А
, О
, У
, Ы
, Э
.
Мягкость
согласного звука обозначается на письме гласными буквами Е
, Ё
, И
, Ю
, Я
, а также буквой Ь
(мягкий знак).
Сравни: нос
[нос] — нёс
[н❜ос], угол
[у́гол] — уголь
[у́гал❜].
Непарные звонкие звуки [й❜], [л], [л❜], [м], [м❜] [н], [н❜] [р], [р❜] называют сонорными
, что в переводе с латинского значит «звучные».
Звуки [ж], [ш], [ч❜], [щ❜] называются шипящими
. Такое название они получили, потому что их произношение похоже на шипение.
Звуки [ж]
, [ш]
— это непарные твёрдые шипящие звуки.
Звуки [ч❜]
и [щ❜]
— это непарные мягкие шипящие звуки.
Звуки [c], [с❜], [з], [з❜], [ц] называются свистящими
.
Согласный звук не бывает
ударным или безударным.
В русском языке согласных звуков (36) больше, чем согласных букв (21), так как одна буква может обозначать парные твёрдый и мягкий звуки: например, буква Л (эль) обозначает звуки [л] и [л❜].
Внимание!
Согласный звук может образовывать слог только с
Основные сведения / Муниципальное казенное общеобразовательное учреждение «Средняя общеобразовательная школа №7»
Директор школы Мананников Евгений Владимирович
«Наша школа — гостеприимный дом талантливых учителей и благодарных учеников»
Муниципальное казенное общеобразовательное учреждение «Cредняя общеобразовательная школа №7» открыла двери перед учащимися в 1989 году, в 2019 году она отметила свой тридцатилетний юбилей.
Свидетельство о государственной регистрации №59244 МКОУ СОШ №7
Филиалов у МКОУ «СОШ №7» — нет.
Постановление от 01.0.2019 года о закреплении муниципальных образовательных организаций за конкретными территориями Октябрьского района на 2019 год
Адрес: 628195, Россия, ХМАО-Югра, Тюменская область, Октябрьский район, г.п.Талинка, 2 мкр., дом №7(здание старшей школы), 1 мкр, дом 50 (здание начальной школы), эл.адрес: [email protected] тел. 8(34672)49915, факс 8(34672)49563
Учредителем школы является Муниципальное образование Октябрьский район представляемое Управлением образования и молодежной политики администрации Октябрьского района, именуемое в дальнейшем Учредитель, расположенное по адресу: ул. Калинина, дом 39, пгт. Октябрьское, Октябрьский район, Тюменская область, Ханты — Мансийский автономный округ — Югра, Россия (почтовый индекс 628100).
Начальник Управления образования -
Киселева Татьяна Борисовна
Сайт:http:/www. oktedu.ru
oktedu.ru
Телефон: 8(34678)28-085
Электронный адрес: [email protected]
Режим и график работы школы
Положение о режиме занятий обучающихся в МКОУ СОШ №7
Календарный график учебного процесса на 2020-2021 (скачать)
Положение о правилах поведения обучающихся МКОУ СОШ №7
Уче6ные занятия в МКОУ «СОШ №7″проводятся в одну смену:
— с 08.00 до 14.00 часов:
— с 14.00 до 21.00 занятия в кружках и секциях.
Учебный год в Школе начинается 1 сентября. Если этот день приходится на выходной день, то учебный год начинается в первый, следующий за ним, рабочий день.
Продолжительность учебного года в Школе: в 1-х классах – 33 учебные недели, в 2-9 классах – 34 учебные недели, 10-11 — 35 учебных недель.
Продолжительность каникул в течение учебного года составляет не менее 30 календарных дней, летом – не менее 8 недель. Для учащихся в первом классе устанавливаются в течение учебного года дополнительные недельные каникулы.
Для учащихся в первом классе устанавливаются в течение учебного года дополнительные недельные каникулы.
Учебный план рассчитан на 5-дневную рабочую неделю во всех классах.
Продолжительность урока 40 минут.
В 1 классе 5-дневная рабочая неделя, продолжительность урока в сентябре, октябре — по 3 урока в день по 35 минут каждый, 1 день 4 урока по 35 минут, в ноябре-декабре — по 4 урока по 35 минут каждый, 1 день 5 уроков по 35 минут; январь — май — по 4 урока по 40 минут каждый, 1 день 5 уроков по 40 минут.
Режим занятий регламентируется календарным учебным графиком.
Недельная образовательная нагрузка (количество учебных занятии) учащихся, реализуемая через урочную и внеурочную деятельность, составляет: 1 класс — 21 час, 2-4 классы — 23 часа, 5 класс — 29 часов, 6 класс – 30 часов, 7 класс – 32 часа, 8 класс – 33 часа, 9-11 классы – 34 часа.
Новости
22.
 04.2021 — Фестиваль национальной кухни
04.2021 — Фестиваль национальной кухни
Праздник национального единства – национальная кухня
Ярким, запоминающимся, хоть и в заочном формате, состоялся традиционный для нашей школы, объединяющий всех «Фес …
15.04.2021 — Встреча участников РДШ МКОУ СОШ №7 с главой администрации гп.Талинка и со специалистами юридического отдела
21 апреля в России отмечается День местного самоуправления. В преддверии этого дня лидеры школьного самоуправления, МКОУ «СОШ №7» из многодетных, малообеспеченных, опекаемых и дети из числа коренных малочисленных народов Севера 14 апреля …
01.04.2021 — Операция «Здоровье»
Операция «Здоровье»
В целях укрепления здоровья, пропаганды здорового образа жизни, в школе ежегодно в плане воспитательной работы школы уделяется особое внимание мероприятиям спортивно-оздоровительного н …
25.03.2021 — Традиционные игры казаков «Шермиции на Оби – 2021»
Традиционные игры казаков «Шермиции на Оби – 2021»
По-разному можно относиться к современному казачеству. Кто-то видит что-то негативное, кто-то выделяет позитивное, но одно не вызывает сомнений – это возможность …
Кто-то видит что-то негативное, кто-то выделяет позитивное, но одно не вызывает сомнений – это возможность …
25.03.2021 — Демонстрационный ЕГЭ-2021 для родителей в МКОУ «СОШ №7»
ЕГЭ-2021 для родителей
25 марта 2021 года к всероссийской акции «Единый день сдачи ЕГЭ родителями» традиционно присоединилась МКОУ «СОШ №7» гп.Талинка. В написа …
Все новости
Сайт школы №19 — г.Рубцовск
Домашнее задание:
ВСЁ ДОМАШНЕЕ ЗАДАНИЕ ДЛЯ 3 Б( ДО 03.02)
РУС.ЯЗ.
1.С 148 ВСЕ СЛОВАРНЫЕ СЛОВА ДО БУКВЫ «З» -УЧИТЬ.
2.с 3-16 все правила!
3.Звуко-буквенный разбор слов: ножка, друг. (ПО ОБРАЗЦУ)
Звуко-буквенный разбор слова
- Согласные звуки – [ Б,В,Г,Д,Ж,З,Й,К,Л,М,Н,П,Р,С,Т,Ф,Х,Ц,Ч,Ш,Щ ]
- Гласные звуки — [А,И,О,У,Ы,Э]
- Буквы Ъ и Ь звука не обозначают
- Всегда мягкие согласные звуки — [Й,Ч,Щ ]
- Всегда твердые согласные звуки — [Ж,Ш,Ц ]
- Звонкие согласные звуки — [ Б,В,Г,Д,Ж,З,Й,Л,М,Н,Р ]
- Глухие согласные звуки — [ К,П,С,Т,Ф,Х,Ц,Ч,Ш,Щ ]
- Парные согласные звуки — [Б-П, В-Ф, З-С, Ж-Ш, Г-К, Д-Т ]
- Гласные буквы Я, Ё, Ю, Я – обозначают два звука:
-в начале слова
-после гласной буквы
-после Ь и Ъ знака
Образец разбора:
лес¹ — [л’эс ] – 1 слог
л [л’ ] – согласный, звонкий, непарный, мягкий
е [э ] – гласный, ударный
с [с ] – согласный, глухой, парный, твердый
_______________________________________
Букв – 3, звуков – 3
Семья¹ — [с’эм’йа ] – 2 слога
с [с’ ] – согласный, глухой, парный, мягкий
е [э ] – гласный, безударный
м [м’ ] – согласный, звонкий, непарный, мягкий
ь –
я [й ] – согласный, звонкий, непарный, мягкий
[а ] – гласный, ударный
__________________________________
Букв – 5, звуков — 5
МАТЕМАТИКА С 6 правило!
С 7№2,№6(если решали решить еще раз ,2 способами и расписать, пример на стр. 6)
6)
с 7 ЗАДАНИЕ ПОД ЧЕРТОЙ, с 8 №6, 7,с10 №4, с 11 задание под чертой.
ОКРУЖАЮЩИЙ МИР С 25-30 читать, с 26-27 сообщение (по выбору), с 30 –подготовиться к опросу на уроке(рубрика «Проверь себя»)
ЧТЕНИЕ -ежедневно читать с записями в дневник по технике чтения
С 47-49 одно стихотворение выучить, другое читать выразительно (по выбору), с 48 и 49-ответить на вопросы.
АГЛИЙСКИЙ ЯЗЫК
1.УЧЕБНИК С 61 № 1 (учить стих и слова в рамке), №4,6 –читать переводить. РТ с 41 №1-4
2.УЧЕБНИК С 63 №3, 5, учить слова в рамке, РТ с 42 №3
28.01
1.рус.яз. с 148 выучить словарные слова на «Б», повторить правило с 5.
тренажер (на повторение): http://igraemsami.ru/russian/glasnye-i-soglasnye-bukvy-onlajn-igra.html(скопируйте ее в адресную строку вашего браузера или нажмите на нее)для улучшения вычислительных навыков, обязательно работайте с ним каждый день!!!
2. матем. повторить таблицу сложения и умножения, вот ссылка на тренировку таблиц сложения и вычитания : http://igraemsami.ru/slozhenie-chisel/slozhenie-igra.html (скопируйте ее в адресную строку вашего браузера или нажмите на нее)для улучшения вычислительных навыков, обязательно работайте с ним каждый день!!!
матем. повторить таблицу сложения и умножения, вот ссылка на тренировку таблиц сложения и вычитания : http://igraemsami.ru/slozhenie-chisel/slozhenie-igra.html (скопируйте ее в адресную строку вашего браузера или нажмите на нее)для улучшения вычислительных навыков, обязательно работайте с ним каждый день!!!
3.чтение РТ с 57-60 выполнить, с 46 выучить стихотворение, с 47 ответить на вопросы.
4.окружающий мир с 14-17 повторить знаки
27.01
1.рус.яз. с 148 выучить словарные слова на «А», повторить правило с 3.
тренажер (на повторение): http://igraemsami.ru/russian/glasnye-i-soglasnye-bukvy-onlajn-igra.html (скопируйте ее в адресную строку вашего браузера или нажмите на нее)для улучшения вычислительных навыков, обязательно работайте с ним каждый день!!!
2.матем. повторить таблицу сложения и умножения, вот ссылка на тренировку таблиц сложения и вычитания : http://igraemsami.ru/slozhenie-chisel/slozhenie-igra.html (скопируйте ее в адресную строку вашего браузера или нажмите на нее)для улучшения вычислительных навыков, обязательно работайте с ним каждый день!!!
3. чтение с 45 познакомиться с разделом, с 45 выучить стихотворение, ознакомиться с биографией поэта:
чтение с 45 познакомиться с разделом, с 45 выучить стихотворение, ознакомиться с биографией поэта:
Черный Саша настоящее имя — Александр Михайлович Гликберг (1880 — 1932), поэт.
Родился 1 октября (13 н.с.) в Одессе в семье провизора. Учился в Петербургской гимназии, из которой был исключен за неуспеваемость. Служил на таможне.
Первые стихи были напечатаны в 1904 в Житомире. В 1905 переехал в Петербург
Впервые подписался псевдонимом «Саша Чёрный» в 1905 под политической сатирой «Чепуха», принесшей ему известность и послужившей поводом для закрытия журнала «Зритель». Первый сборник стихов «Разные мотивы», где наряду с лирикой были политические сатиры, был запрещен цензурой.
В 1906 — 1907 живет в Германии. Вернувшись в Петербург, в течение трех лет (1908 — 11) сотрудничает с «Сатириконом» (журнал сатиры и юмора), становится ведущим автором, приобретает всероссийскую известность. Уйдя из «Сатирикона», писал для различных газет и журналов, написал несколько детских книжек («Тук-тук», 1913; «Живая азбука», 1914), занимался переводами.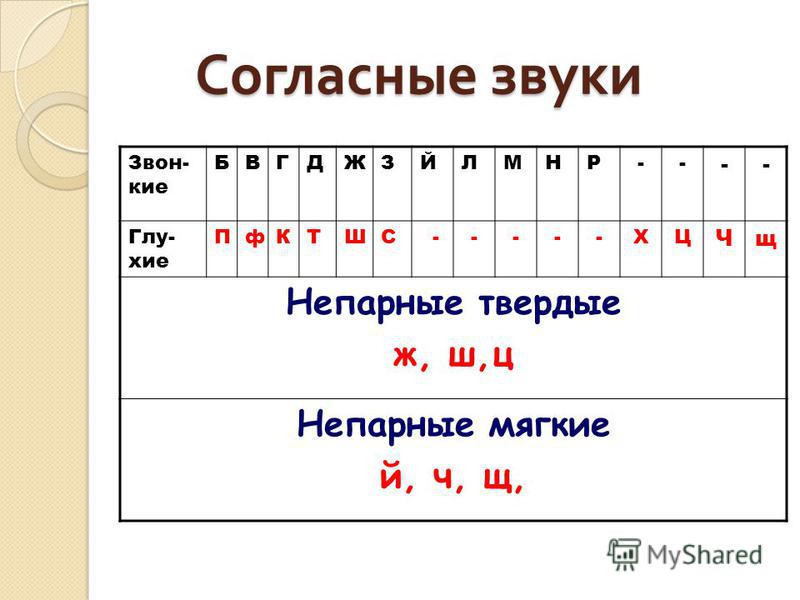
Написал цикл стихов для детей. жил в Германии, выпустив третью книгу стихов — «Жажда» (1923). Выступал в основном как детский писатель.
В последние годы в его творчестве большое место занимала проза («Солдатские сказки», опубликованные в 1933; «Несерьезные рассказы», 1928).
Умер Саша Черный во Франции, в местечке Лаванду, 5 июля 1932.
Непарные твердые согласные звуки. Парные и непарные, звонкие и глухие, мягкие и твердые согласные звуки в русском языке
Многие русские согласные образуют пары по твердости-мягкости: – , – и другие. Звуки, соответствующие ударным и, после мягких согласных в слабой, безударной позиции звучат одинаково. Буква обозначает звук, например гласные после твёрдых согласных и согласные перед гласными: погода.
Ведущая Василиса попросила повторить всё, что выучили ученики про согласные. Друзья из Шишкиного Леса вспомнили немало: Согласных больше чем гласных. Согласные нельзя спеть. Они произносятся с шумом и голосом: Б, Ж, З. Или только с шумом: П, Т, Ф. Согласные бывают звонкие глухие парные непарные.
Они произносятся с шумом и голосом: Б, Ж, З. Или только с шумом: П, Т, Ф. Согласные бывают звонкие глухие парные непарные.
§6. Твёрдые и мягкие согласные
Всё дело в том, что ты пропустил предыдущий урок, на котором мы изучали парные согласные, — пояснила Василиса. Звонкому «Ж» парным будет глухой «Ш». Например: жар — шар. — Я понял, – сказал Зубок. Глухой звук это то же что и звонкий, но сказанный тихо, без голоса. Достаточно изменить слово так, чтобы после непонятного согласного стояла гласная. Однако не все согласные парные.
В одном будут жить парные согласные, а в другом — непарные. Парные Непарные Ж — Ш М, Н З — С Х, Ц К — Г Р, Л А теперь давайте составим рассказ из слов, в которых только непарные согласные. Хоть и парные эти согласные, Но они всё равно очень разные. В безударной позиции гласные произносятся менее четко и звучат с меньшей длительностью (т.е. редуцируются).
Сколько пар образуют согласные по глухости-звонкости?
Не забывайте, что парные звонкие согласные в слабой позиции на конце слова или перед глухим согласным всегда оглушаются, а глухие перед звонким – иногда озвончаются. Когда буквы, обыкновенно обозначающие глухие согласные, при озвончении обозначают звонкие звуки, это кажется настолько необычным, что может привести к ошибкам в транскрипции. В заданиях, связанных со сравнением количества букв и звуков в слове, могут быть «ловушки», провоцирующие ошибки.
Когда буквы, обыкновенно обозначающие глухие согласные, при озвончении обозначают звонкие звуки, это кажется настолько необычным, что может привести к ошибкам в транскрипции. В заданиях, связанных со сравнением количества букв и звуков в слове, могут быть «ловушки», провоцирующие ошибки.
Возможны слова, которые могут состоять только из гласных, но согласные тоже необходимы. В русском языке согласных намного больше, чем гласных. Согласные – это звуки, при произнесении которых воздух встречает на своём пути преграду. В русском языке два вида преграды: щель и смычка – это два основных способа образования согласных.
Смычка, второй вид артикуляции согласных,образуется при смыкании органов речи. Поток воздуха резко преодолевает эту преграду, звуки получаются краткими, энергичными. Сравним слова: дом и кот. В каждом слове по 1-му гласному звуку и по 2 согласных.
2) перед ними не происходит озвончение парных глухих согласных (т.е. позиция перед ними сильная по глухости-звонкости, как и перед гласными).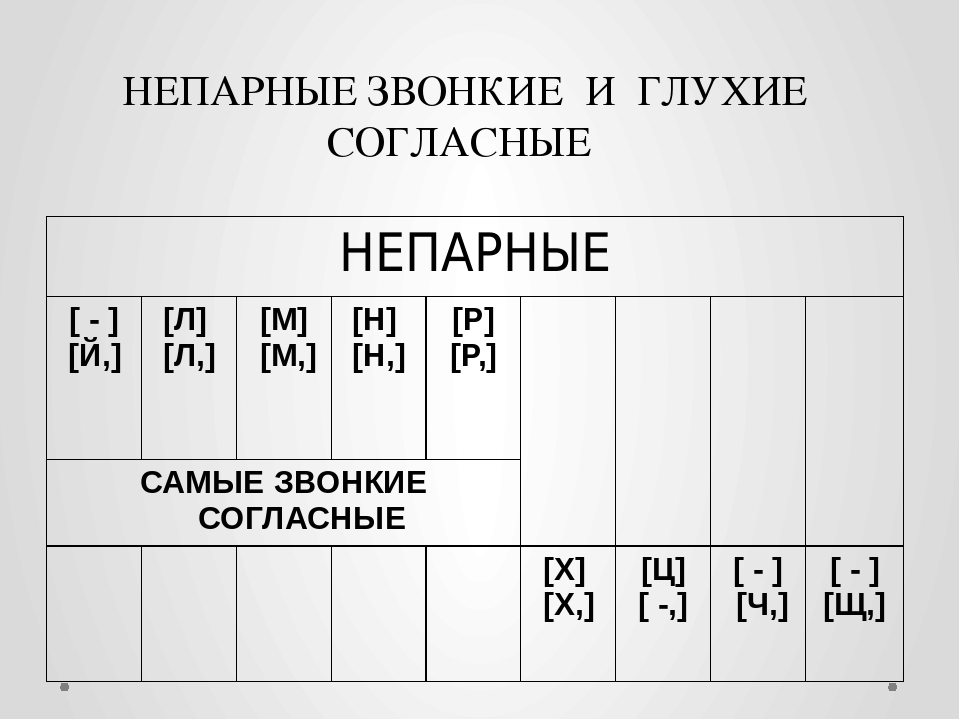 Но есть звуки, у которых нет пары по признаку твёрдости-мягкости. В школьных учебниках сказано, что и — непарные по твёрдости-мягкости. Как же так? Мы ведь слышим, что звук – это мягкий аналог звука.Когда в школе училась я сама, я не могла понять почему?
Но есть звуки, у которых нет пары по признаку твёрдости-мягкости. В школьных учебниках сказано, что и — непарные по твёрдости-мягкости. Как же так? Мы ведь слышим, что звук – это мягкий аналог звука.Когда в школе училась я сама, я не могла понять почему?
Парные по звонкости-глухости согласные звуки
Недоумение возникает, потому что школьные учебники не учитывают, что звук ещё и долгий, а твёрдый нет. Пары – это звуки, различающиеся только одним признаком. А и – двумя. Поэтому и не являются парами. Во-первых, ребята поначалу часто смешивают звуки и буквы. Использование буквы в транскрипции создаст основание для такого смешения, спровоцирует ошибку.
Нужно понять, осмыслить, а потом запомнить, что на самом деле звуки и пару по твёрдости-мягкости не образуют. Важны условия, в которых оказывается тот или иной звук. Начало слова, конец слова, ударный слог, безударный слог, положение перед гласным, положение перед согласным – всё это разные позиции.
В безударных слогах гласные подвергаются изменениям: они короче и не произносятся так же отчётливо, как под ударением. И под ударением, и в безударной позиции мы ясно слышим: , и пишем буквы, которыми эти звуки принято обозначать. Упростили. Но многие ребята с хорошим слухом, слышащие ясно, что звуки в следующих примерах разные, никак не могут понять, почему учитель и учебник настаивают на том, что эти звуки одинаковы.
И под ударением, и в безударной позиции мы ясно слышим: , и пишем буквы, которыми эти звуки принято обозначать. Упростили. Но многие ребята с хорошим слухом, слышащие ясно, что звуки в следующих примерах разные, никак не могут понять, почему учитель и учебник настаивают на том, что эти звуки одинаковы.
В ней наблюдается прояснение гласных после мягких согласных. Позиционные изменения наблюдаются только у парных согласных. Во всех случаях в слабой позиции возможно позиционное смягчение согласных. Естественно, в школьной традиции не принято излагать характеристики звуков и происходящих с ними позиционных изменений со всеми подробностями. Поэтому ниже представлен список позиционно-обусловленных изменений согласных по признакам способа и места образования.
Буква может обозначать качество предшествующего звука, например ь в словах тень, пень, пальба. Сравнение с гласными звуками. Каждый согласный имеет признаки, отличающие его от остальных согласных звуков. В речи может происходить замена звуков под влиянием соседних звуков в слове. Важно знать сильные и слабые позиции согласных звуков в слове для их правильного написания.
Важно знать сильные и слабые позиции согласных звуков в слове для их правильного написания.
Классификация согласных.
Если человек произносит согласные звуки, то закрывает (хоть немного) рот, из-за этого получается шум. Но шумят согласные по-разному. Заселим фонетические домики в городе звуков. Договоримся: на первом этаже будут жить глухие звуки, а на втором – звонкие.
У звуков и нет парных мягких звуков, они всегда твёрдые. Но не все согласные звуки и буквы образуют пары. Те согласные, которые пар не имеют, называют непарные. Поселим непарные согласные звуки в свои домики. Звуки второго домика называют ещё сонорными, потому что образуются они с помощью голоса и почти без шума, они очень звучные. В первую положим те, в названиях которых слышно какие-либо мягкие звуки, во вторую те, в названиях которых все согласные звуки твёрдые.
Чтобы не перепутать при прочтении транскрипции твёрдые и мягкие звуки, учёные договорились показывать мягкость звука значком, очень похожим на запятую, только ставят его сверху.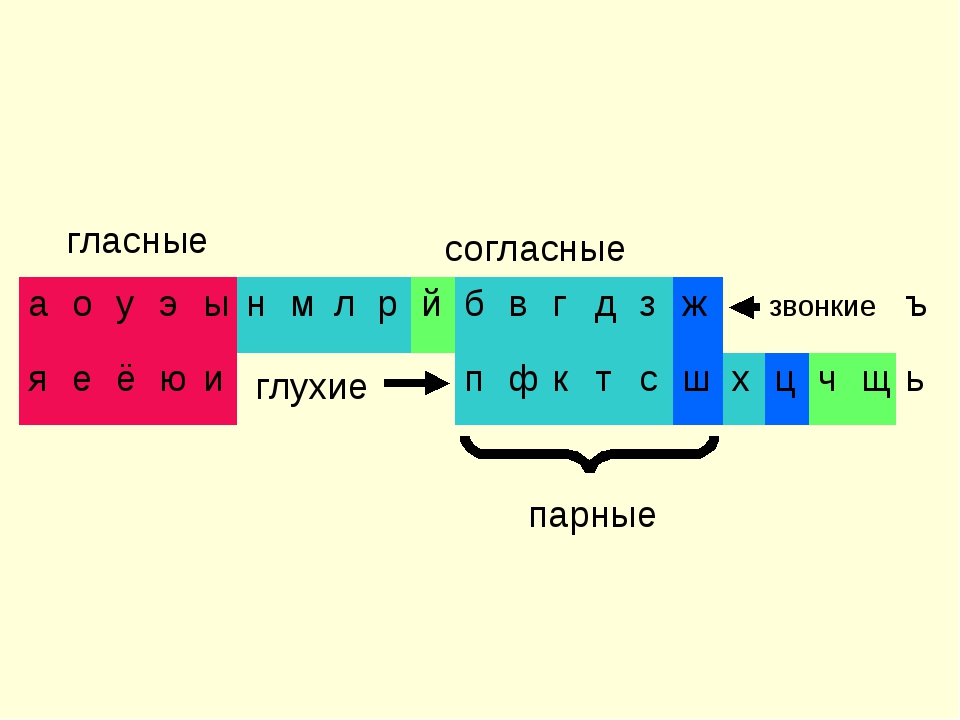
И тогда мы поймём точно — какую букву нужно писать. Давайте вместе найдём в русской азбуке этих одиночек. Он этого не заметил, потому что смотрел на луну. И тут вошёл его верный рыцарь. И спугнул муху. Молодцы! То ли звонкий, то ли потише, Кот — кота, год — года. Различим без труда. И в конец букву верно напишем. Гласные, без ударения в целом сохраняют свое звучание. Буквы е, ё, ю, я играют двойную роль в русской графике. Звук – минимальная единица звучащей речи. У каждого слова есть звуковая оболочка, состоящая из звуков.
Звуки делятся на гласные и согласные.У них разная природа. По соотношению шума и голоса согласные делятся на звонкие и глухие. Нормативным произношением считается «иканье», т.е. неразличение Э и А в безударном положении после мягких согласных. Такое изменение гласных в слабой позиции называется редукцией. В слове гласные могут быть в ударных и безударных слогах. В слабых позициях согласные видоизменяются: с ними происходят позиционные изменения.
Звуком называется мельчайшая единица языка, произносимая с помощью органов речевого аппарата. Ученые обнаружили, что при рождении человеческий слух воспринимает все звуки, которые слышит. Все это время его мозг отсортировывает ненужную информацию, и уже к 8-10 месяцам человек способен различать звуки, присущие исключительно родному языку, и все нюансы произношения.
Ученые обнаружили, что при рождении человеческий слух воспринимает все звуки, которые слышит. Все это время его мозг отсортировывает ненужную информацию, и уже к 8-10 месяцам человек способен различать звуки, присущие исключительно родному языку, и все нюансы произношения.
33 буквы составляют русский алфавит, 21 из них являются согласными, однако следует отличать буквы от звуков. Буквой является знак, символ, который можно увидеть или написать. Звук можно только услышать и произнести, а на письме — обозначить с помощью транскрипции — [б], [в], [г]. Они несут в себе определенную смысловую нагрузку, соединяясь между собой, образуют слова.
36 согласных звуков: [б], [з], [в], [д], [г], [ж], [м], [н], [к], [л], [т], [п], [т], [с], [щ], [ф], [ц], [ш], [х], [ч], [б»], [з»], [в»], [д»], [й»], [н»], [к»], [м»], [л»], [т»], [с»], [п»], [р»], [ф»], [г»], [х»].
Согласные звуки делятся на:
- мягкие и твердые;
звонкие и глухие;
парные и непарные.
Мягкие и твердые согласные звуки
Фонетика русского языка имеет существенное отличие от многих других языков. Она содержит твердые и мягкие согласные.
Она содержит твердые и мягкие согласные.
В момент произношения мягкого звука язык сильнее прижимается к небу, чем при произнесении твердого согласного звука, препятствуя высвобождению воздуха. Этим и отличается друг от друга твердый и мягкий согласный звук. Для того чтобы на письме определить, относится ли согласный звук к мягким или твердым, следует посмотреть на букву, стоящую сразу после конкретной согласной.
Согласные звуки относят к твердым в таких случаях:
- если буквы а, о, у, э, ы
следуют после них — [мак], [ром], [гул], [сок], [бык]; - после них стоит другой согласный звук — [ворс], [град], [брак];
- если звук стоит в конце слова — [мрак], [друг], [стол].
Мягкость звука записывается в виде апострофа: моль — [мол’], мел — [м’эл], калитка — [кал’итка], пир — [п’ир].
Следует отметить, что звуки [щ’], [й’], [ч’] всегда мягкие, и твердые согласные — только [ш], [ц], [ж].
Согласный звук станем мягким, если после него стоит «ь» и гласные: я, ё, ю, и, е.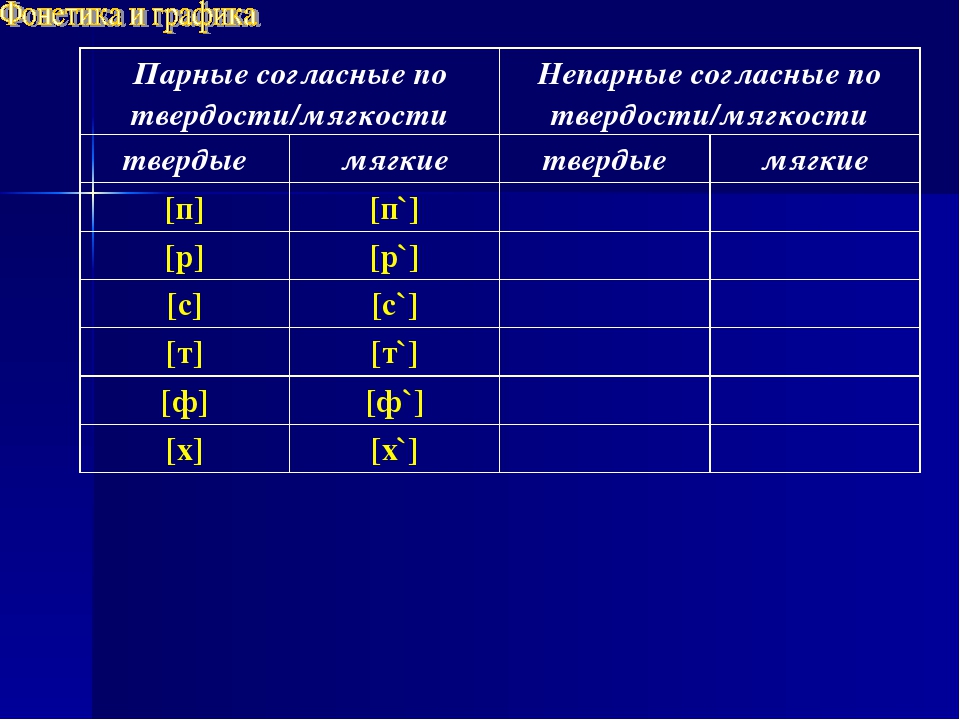 Например: ген — [г»эн], лён — [л»он], диск — [д»ыск], люк — [л»ук], вяз — [в»яз], трель — [тр»эл»].
Например: ген — [г»эн], лён — [л»он], диск — [д»ыск], люк — [л»ук], вяз — [в»яз], трель — [тр»эл»].
Звонкие и глухие, парные и непарные звуки
По звонкости согласные разделяют на звонкие и глухие. Звонкими согласными могут быть звуки, создаваемые с участием голоса: [в], [з], [ж], [б], [г], [й], [м], [д], [л], [р], [н].
Примеры: [бор], [вол], [душ], [зов], [жар], [гол], [лов], [мор], [нос], [род], [рой].
Примеры: [кол], [пол], [том], [сон], [шум], [щ»ука], [хор], [цар»], [ч»ан].
К парным звонким и глухим согласным относятся: [б] — [п], [ж] — [ш] , [г] — [х], [з] — [с]. [д] — [т], [в] — [ф]. Примеры: быль — пыль, дом — том, год — код, ваза — фаза, зуд — суд, жить — шить.
Звуки, не образующие пары: [ч], [н], [ц], [х], [р], [м], [л].
Мягкие и твердые согласные тоже могут иметь пару: [р] — [р»], [п] — [п»], [м] — [м»], [в] — [в»], [д] — [д»], [ф] — [ф»], [к] — [к»], [з] — [з»], [б] — [б»], [г] — [г»], [н] — [н»], [с] — [с»], [л] — [л»], [т] — [т»], [х] — [х»].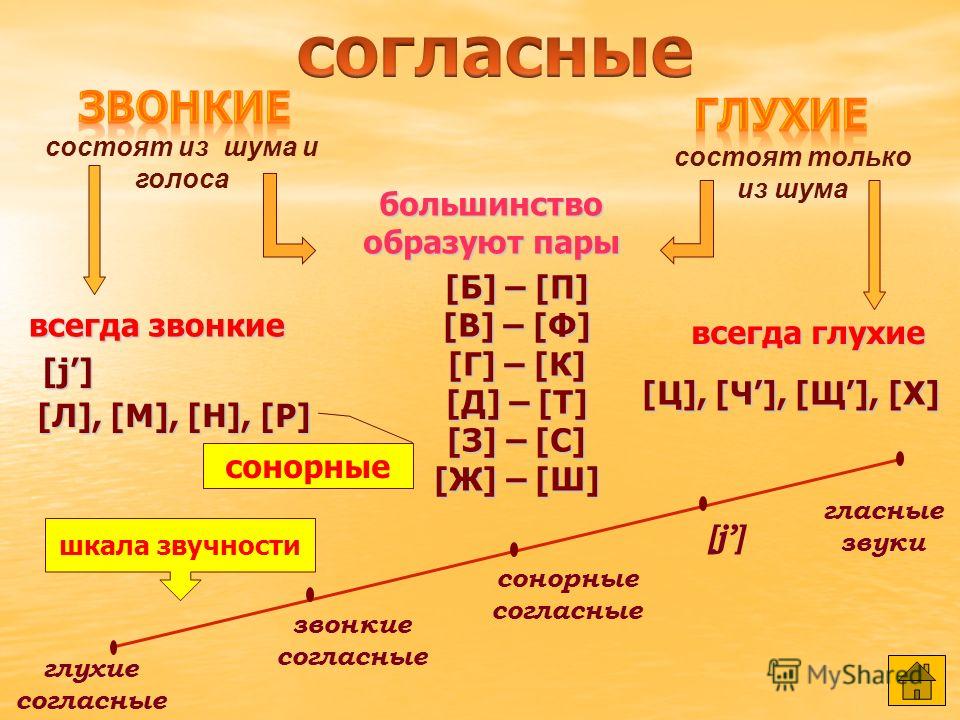 Примеры: быль — бель, высь — ветвь, город — гепард, дача — дело, зонт — зебра, кожа — кедр, луна — лето, монстр — место, палец — перо, руда — река, сода — сера, столб — степь, фонарь — ферма, хоромы — хижина.
Примеры: быль — бель, высь — ветвь, город — гепард, дача — дело, зонт — зебра, кожа — кедр, луна — лето, монстр — место, палец — перо, руда — река, сода — сера, столб — степь, фонарь — ферма, хоромы — хижина.
Таблица для запоминания согласных звуков
Чтобы наглядно увидеть и сравнить мягкие и твердые согласные, таблица, приведенная ниже, их показывает попарно.
Твердые — перед буквами А, О, У, Ы, Э Мягкие — перед буквами И, Е, Ё, Ю, Я | Твердые и мягкие согласные | |||
| б | бал | б» | битва | |
| в | вой | в» | веко | |
| г | гараж | г» | герой | |
| д | дыра | д» | дёготь | |
| з | зола | з» | зевок | |
| к | кум | к» | кеды | |
| л | лоза | л» | листва | |
| м | март | м» | месяц | |
| н | нога | н» | нежность | |
| п | паук | п» | песня | |
| р | рост | р» | ревень | |
| с | соль | с» | сено | |
| т | туча | т» | терпение | |
| ф | фосфор | ф» | фирма | |
| х | худоба | х» | химия | |
| Непарные | ж | жираф | ч | чудо |
| ш | ширма | щ | лещина | |
| ц | цель | й | войлок | |
Запомнить согласные звуки поможет и другая таблица.
| Парные | Звонкие | Глухие |
| Б | П | |
| В | Ф | |
| Г | К | |
| Д | Т | |
| Ж | Ш | |
| З | С | |
| Непарные | Л, М, Н, Р, Й | Х, Ц, Ч, Щ |
Детские стихи для лучшего освоения материала
Буквы ровно 33 в русском алфавите,
Чтоб узнать, сколько согласных —
Десять гласных отнимите,
Знаки — твердый, мягкий —
Сразу станет ясно:
Получается число ровно двадцать одно.
Мягкие и твердые согласные бывают очень разные,
Но вовсе не опасные.
Если произносим с шумом, то они глухие.
Звуки согласные гордо говорят:
Они по-разному звучат.
Твердые и мягкие
На деле очень легкие.
Одно простое правило запомни навсегда:
Ш, Ц, Ж — тверды всегда,
А вот Ч, Щ, Й — только мягки,
Как кошачьи лапки.
А другие смягчим так:
Если добавим мягкий знак,
Тогда получим ель, моль, соль,
Какой же хитрый знак такой!
А если мы добавим гласные И, Я, Ё, Е, Ю,
Получим мягкую согласную.
Знаки-братья, мягкий, твердый,
Мы не произносим,
Но чтоб слово изменить,
Их помощи попросим.
Всадник скачет на коне,
Кон — используем в игре.
На этом уроке мы научимся различать звонкие и глухие согласные звуки и обозначать их на письме согласными буквами. Узнаем, какие согласные называют парными и непарными по звонкости — глухости, сонорными и шипящими.
Звонкие и глухие согласные звуки
Вспомним, как рождаются звуки речи. Когда человек начинает говорить, то выдыхает воздух из лёгких. По дыхательному горлу он бежит в узенькую гортань, где находятся особые мышцы — голосовые связки. Если человек произносит согласные звуки, то закрывает (хоть немного) рот, из-за этого получается шум. Но шумят согласные по-разному.
Когда человек начинает говорить, то выдыхает воздух из лёгких. По дыхательному горлу он бежит в узенькую гортань, где находятся особые мышцы — голосовые связки. Если человек произносит согласные звуки, то закрывает (хоть немного) рот, из-за этого получается шум. Но шумят согласные по-разному.
Проведем опыт: зажмем уши и произнесем звук [п], а потом звук [б]. Когда мы произнесли звук [б], связки натянулись и начали дрожать. Это дрожание превратилось в голос. В ушах немного зазвенело.
Можно провести похожий опыт, положив руки на шею с правой и левой стороны, и произнести звуки [д] и [т]. Звук [д] произносится намного звонче, более звучно. Такие звуки учёные назвали звонкими
, а звуки, которые состоят только из шума — глухими.
Парные по звонкости-глухости согласные звуки
Попробуем разделить звуки на две группы по способу произношения. Заселим фонетические домики в городе звуков. Договоримся: на первом этаже будут жить глухие звуки, а на втором — звонкие. Жители первого домика:
Жители первого домика:
Эти согласные звуки называются парными
по звонкости — глухости.
Рис. 1. Парные звонкие и глухие согласные звуки ()
Они очень похожи друг на друга — настоящие «двойняшки», их и произносят почти одинаково: одинаково складываются губы, одинаково двигается язык. Но у них есть пары и по мягкости — твёрдости. Добавим их в домик.
У звуков [ж] и [ш] нет парных мягких звуков, они всегда твёрдые
. А ещё их называют шипящими
звуками.
Все эти звуки обозначаются буквами:
Непарные звонкие согласные звуки
Но не все согласные звуки и буквы образуют пары. Те согласные, которые пар не имеют, называют непарные.
Поселим непарные согласные звуки в свои домики.
Во второй домик — непарные
звонкие согласные
звуки:
Напомним, что звук [й’] всегда только мягкий.
Поэтому в нашем домике он будет жить один. Эти звуки обозначают на письме буквами:
Звуки второго домика называют ещё сонорными
, потому что образуются они с помощью голоса и почти без шума, они очень звучные.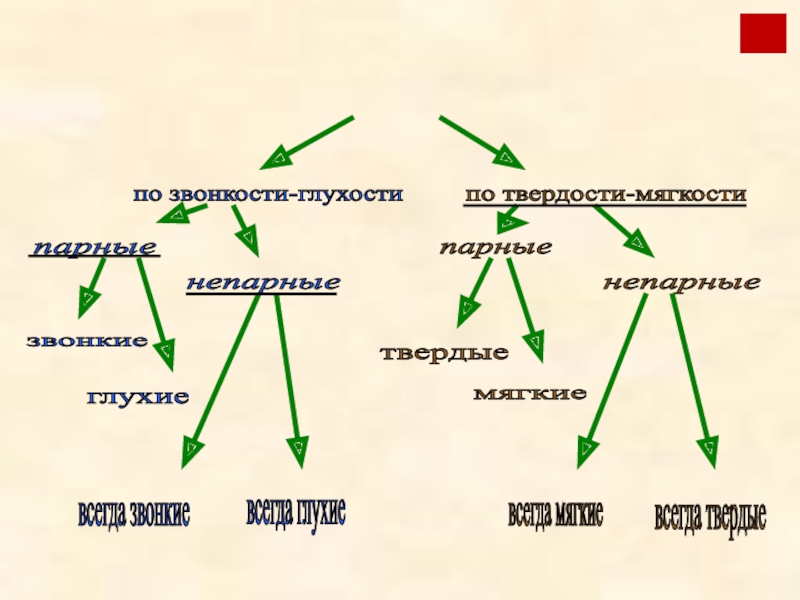 Слово «сонорный» в переводе с латинского «сонорус» означает звонкий.
Слово «сонорный» в переводе с латинского «сонорус» означает звонкий.
Непарные глухие согласные звуки
В третий домик мы поселим непарные глухие согласные
звуки:
Вспомним, что звук [ц] всегда твёрдый
, а [ч’] и [щ’] — всегда мягкие.
Непарные глухие согласные звуки обозначают на письме буквами:
Звуки [ч’], [щ’] — шипящие
звуки.
Вот мы и заселили наш город согласных звуков и букв. Теперь сразу понятно, почему согласных букв — 21, а звуков — 36.
Рис. 2. Звонкие и глухие согласные звуки ()
Закрепление знаний на практике
Выполним задания.
1. Рассмотрим картинки и превратим одно слово в другое, заменив только один звук. Подсказка: вспомним пары согласных звуков.
д
очка — точка
б
очка — почка
ш
ар — жар
удочка — уточка
2. Существуют загадки, смысл которых заключён в знании согласных звуков, они называются шарады.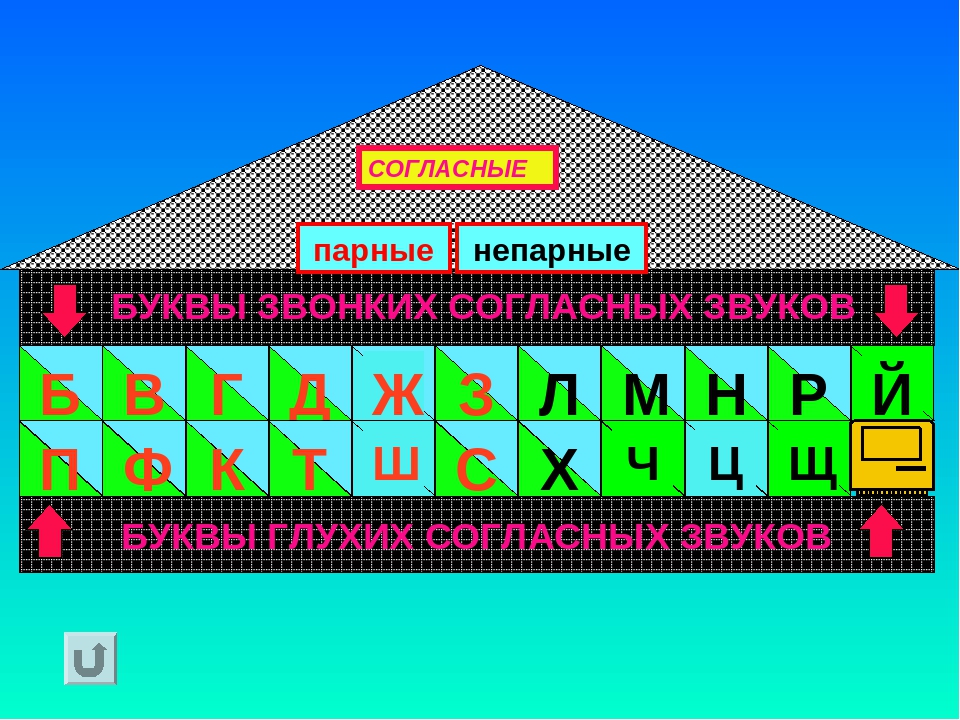 Попробуйте их отгадать:
Попробуйте их отгадать:
1) С глухим согласным наливаюсь в поле,
Со звонким — сам звеню я на раздолье. (Колос — голос
)
2) С глухим — траву она срезает,
Со звонким — листья объедает. (Коса — коза)
3) С «эм» — приятен, золотист, очень сладок и душист.
С буквой «эль» зимой бывает, а весною исчезает. (Мёд- лёд)
Для того чтобы развить умение произносить какие-то звуки, особенно шипящие, учат скороговорки. Скороговорку рассказывают сначала медленно, а потом ускоряют темп. Попробуем выучить скороговорки:
- Шесть мышат в камышах шуршат.
- У ежа — ежата, у ужа — ужата.
- Два щенка щека к щеке грызли щётку в уголке.
Итак, сегодня мы узнали, что согласные звуки могут быть звонкими и глухими и как обозначаются на письме эти звуки.
- Андрианова Т.М., Илюхина В.А. Русский язык 1. М.: Астрель, 2011. ().
- Бунеев Р.Н., Бунеева Е.В., Пронина О.
 В. Русский язык 1. М.: Баллас. ().
В. Русский язык 1. М.: Баллас. (). - Агаркова Н.Г., Агарков Ю.А. Учебник по обучению грамоте и чтению: Азбука. Академкнига/Учебник.
- Fictionbook.ru ().
- Deafnet.ru ().
- Samouchka.com.ua ().
- Андрианова Т.М., Илюхина В.А. Русский язык 1. М.: Астрель, 2011. Стр. 38, упр. 2; Стр. 39, упр. 6; Стр. 43, упр. 4.
- Сосчитайте, сколько звонких согласных и сколько глухих в слове неудовлетворительный
?
(Звонких согласных — 9 — Н, Д, В, Л, В, Р, Л, Н, Й, различных -6, глухих согласных — 2 — Т, Т, различных — 1.)
. - Прочитайте пословицу: «
Умей вовремя сказать, вовремя смолчать».
Назовите буквы, которые обозначают звонкие согласные звуки. (Звонкие согласные звуки обозначают в пословице буквы М, Й, В, Р, З, Л.)
- 4* Используя полученные на уроке знания, напишите сказку или нарисуйте комикс на тему «В городе согласных звуков».
На этом уроке мы научимся различать звонкие и глухие согласные звуки и обозначать их на письме согласными буквами. Узнаем, какие согласные называют парными и непарными по звонкости — глухости, сонорными и шипящими.
Узнаем, какие согласные называют парными и непарными по звонкости — глухости, сонорными и шипящими.
Звонкие и глухие согласные звуки
Вспомним, как рождаются звуки речи. Когда человек начинает говорить, то выдыхает воздух из лёгких. По дыхательному горлу он бежит в узенькую гортань, где находятся особые мышцы — голосовые связки. Если человек произносит согласные звуки, то закрывает (хоть немного) рот, из-за этого получается шум. Но шумят согласные по-разному.
Проведем опыт: зажмем уши и произнесем звук [п], а потом звук [б]. Когда мы произнесли звук [б], связки натянулись и начали дрожать. Это дрожание превратилось в голос. В ушах немного зазвенело.
Можно провести похожий опыт, положив руки на шею с правой и левой стороны, и произнести звуки [д] и [т]. Звук [д] произносится намного звонче, более звучно. Такие звуки учёные назвали звонкими
, а звуки, которые состоят только из шума — глухими.
Парные по звонкости-глухости согласные звуки
Попробуем разделить звуки на две группы по способу произношения. Заселим фонетические домики в городе звуков. Договоримся: на первом этаже будут жить глухие звуки, а на втором — звонкие. Жители первого домика:
Заселим фонетические домики в городе звуков. Договоримся: на первом этаже будут жить глухие звуки, а на втором — звонкие. Жители первого домика:
Эти согласные звуки называются парными
по звонкости — глухости.
Рис. 1. Парные звонкие и глухие согласные звуки ()
Они очень похожи друг на друга — настоящие «двойняшки», их и произносят почти одинаково: одинаково складываются губы, одинаково двигается язык. Но у них есть пары и по мягкости — твёрдости. Добавим их в домик.
У звуков [ж] и [ш] нет парных мягких звуков, они всегда твёрдые
. А ещё их называют шипящими
звуками.
Все эти звуки обозначаются буквами:
Непарные звонкие согласные звуки
Но не все согласные звуки и буквы образуют пары. Те согласные, которые пар не имеют, называют непарные.
Поселим непарные согласные звуки в свои домики.
Во второй домик — непарные
звонкие согласные
звуки:
Напомним, что звук [й’] всегда только мягкий.
Поэтому в нашем домике он будет жить один. Эти звуки обозначают на письме буквами:
Звуки второго домика называют ещё сонорными
, потому что образуются они с помощью голоса и почти без шума, они очень звучные. Слово «сонорный» в переводе с латинского «сонорус» означает звонкий.
Непарные глухие согласные звуки
В третий домик мы поселим непарные глухие согласные
звуки:
Вспомним, что звук [ц] всегда твёрдый
, а [ч’] и [щ’] — всегда мягкие.
Непарные глухие согласные звуки обозначают на письме буквами:
Звуки [ч’], [щ’] — шипящие
звуки.
Вот мы и заселили наш город согласных звуков и букв. Теперь сразу понятно, почему согласных букв — 21, а звуков — 36.
Рис. 2. Звонкие и глухие согласные звуки ()
Закрепление знаний на практике
Выполним задания.
1. Рассмотрим картинки и превратим одно слово в другое, заменив только один звук. Подсказка: вспомним пары согласных звуков.
д
очка — точка
б
очка — почка
ш
ар — жар
удочка — уточка
2. Существуют загадки, смысл которых заключён в знании согласных звуков, они называются шарады. Попробуйте их отгадать:
1) С глухим согласным наливаюсь в поле,
Со звонким — сам звеню я на раздолье. (Колос — голос
)
2) С глухим — траву она срезает,
Со звонким — листья объедает. (Коса — коза)
3) С «эм» — приятен, золотист, очень сладок и душист.
С буквой «эль» зимой бывает, а весною исчезает. (Мёд- лёд)
Для того чтобы развить умение произносить какие-то звуки, особенно шипящие, учат скороговорки. Скороговорку рассказывают сначала медленно, а потом ускоряют темп. Попробуем выучить скороговорки:
- Шесть мышат в камышах шуршат.
- У ежа — ежата, у ужа — ужата.
- Два щенка щека к щеке грызли щётку в уголке.
Итак, сегодня мы узнали, что согласные звуки могут быть звонкими и глухими и как обозначаются на письме эти звуки.
- Андрианова Т.М., Илюхина В.А. Русский язык 1. М.: Астрель, 2011. ().
- Бунеев Р.Н., Бунеева Е.В., Пронина О.В. Русский язык 1. М.: Баллас. ().
- Агаркова Н.Г., Агарков Ю.А. Учебник по обучению грамоте и чтению: Азбука. Академкнига/Учебник.
- Fictionbook.ru ().
- Deafnet.ru ().
- Samouchka.com.ua ().
- Андрианова Т.М., Илюхина В.А. Русский язык 1. М.: Астрель, 2011. Стр. 38, упр. 2; Стр. 39, упр. 6; Стр. 43, упр. 4.
- Сосчитайте, сколько звонких согласных и сколько глухих в слове неудовлетворительный
?
(Звонких согласных — 9 — Н, Д, В, Л, В, Р, Л, Н, Й, различных -6, глухих согласных — 2 — Т, Т, различных — 1.)
. - Прочитайте пословицу: «
Умей вовремя сказать, вовремя смолчать».
Назовите буквы, которые обозначают звонкие согласные звуки. (Звонкие согласные звуки обозначают в пословице буквы М, Й, В, Р, З, Л.)
- 4* Используя полученные на уроке знания, напишите сказку или нарисуйте комикс на тему «В городе согласных звуков».
Звонкие согласные таблица. Русский язык — Звонкие и глухие, парные и непарные согласные звуки — видео — телеканал «Радость моя. Сильные-слабые позиции для гласных. Позиционные изменения гласных. Редукция
В русском языке 21 согласная буква и 36 согласных звуков. Согласные буквы и соответствующие им согласные звуки:
б — [б], в — [в], г — [г], д — [д], ж — [ж], й — [й], з — [з], к — [к], л — [л], м — [м], н — [н], п — [п], р — [р], с — [с], т — [т], ф — [ф], х — [х], ц — [ц], ч — [ч], ш — [ш], щ — [щ].
Согласные звуки делятся на звонкие и глухие, твёрдые и мягкие. Они бывают парные и непарные. Всего 36 различных комбинаций согласных по парности-непарности твёрдых и мягких, глухих и звонких: глухих — 16 (8 мягких и 8 твёрдых), звонких — 20 (10 мягких и 10 твёрдых).
Схема 1. Согласные буквы и согласные звуки русского языка.
Твёрдые и мягкие согласные звуки
Согласные бывают твёрдыми и мягкими. Они делятся на парные и непарные. Парные твёрдые и парные мягкие согласные помогают нам различать слова. Сравните: конь [кон’] — кон [кон], лук [лук] — люк [л’ук].
Для понимания объясним «на пальцах». Если согласная буква в разных словах означает либо мягкий, либо твёрдый звук, то звук относится к парным. Например, в слове кот буква к обозначает твёрдый звук [к], в слове кит буква к обозначает мягкий звук [к’]. Получаем: [к]-[к’] образуют пару по твёрдости-мягкости. Нельзя относить к паре звуки для разных согласных, например [в] и [к’] не составляют пару по твёрдости-мягкости, но составляет пара [в]-[в’]. Если согласный звук всегда твёрдый либо всегда мягкий, то он относится к непарным согласным. Например, звук [ж] всегда твёрдый. В русском языке нет слов, где бы он был мягким [ж’]. Так как не бывает пары [ж]-[ж’], то он относится к непарным.
Звонкие и глухие согласные звуки
Согласные звуки бывают звонкие и глухие. Благодаря звонким и глухим согласным мы различаем слова. Сравните: шар — жар, кол — гол, дом — том. Глухие согласные произносятся почти с прикрытым ртом, при их произнесении голосовые связки не работают. Для звонких согласных нужно больше воздуха, работают голосовые связки.
Некоторые согласные звуки имеют схожее звучание по способу произношения, но произносятся с разной тональностью — глухо или звонко. Такие звуки объединяются в пары и образуют группу парных согласных. Соответственно, парные согласные — это пара из глухой и звонкой согласной.
- парные согласные: б-п, в-ф, г-к, д-т, з-с, ж-ш.
- непарные согласные: л, м, н, р, й, ц, х, ч, щ.
Сонорные, шумные и шипящие согласные
Сонорные — звонкие непарные согласные звуки. Сонорных звуков 9: [й’], [л], [л’], [м], [м’], [н], [н’], [р], [р’].
Шумные согласные звуки бывают звонкие и глухие:
- Шумные глухие согласные звуки (16): [к], [к»], [п], [п»], [с], [с»], [т], [т»], [ф], [ф»], [х], [х’], [ц], [ч’], [ш], [щ’];
- Шумные звонкие согласные звуки (11): [б], [б’], [в], [в’], [г], [г’], [д], [д’], [ж], [з], [з’].
Шипящие согласные звуки (4): [ж], [ч’], [ш], [щ’].
Парные и непарные согласные звуки
Согласные звуки (мягкие и твёрдые, глухие и звонкие) делятся на парные и непарные. Выше в таблицах показано деление. Обобщим всё схемой:
Схема 2. Парные и непарные согласные звуки.
Чтобы уметь делать фонетический разбор, помимо согласных звуков нужно знать
Как известно, звуки речи можно разделить на гласные (произносимые только голосом) и согласные (в их произнесении участвует шум). Многие согласные можно разбить на пары по их характеристикам, но не все.
Парные и непарные согласные по глухости-звонкости
Сразу надо оговориться, что таких звуков, которые непарные по всем признакам, всего четыре. Мы поговорим о них в конце статьи. Большинство же по одному признаку входит в пару, а по другому — нет. Поэтому бессмысленно писать про согласный «непарный» – надо указать, по какому признаку.
Согласные различаются по глухости-звонкости. Это означает, что при произнесении одних из них используется больше голоса (сонорные, звонкие), а в других больше шума (глухие) или даже вообще один шум (шипящие).
Сонорные — это очень звонкие согласные, голоса в них много, а шума мало.
Два сонорных согласных — [Л] и [Р] – даже могут при некоторых обстоятельствах образовывать слог, то есть вести себя как гласные. Наверняка вы встречали ошибочное написание «театор». Оно объясняется как раз тем, что [Р] в этом слове слогообразующее. Другие примеры — слова «Александр», «смысл».
Непарные звонкие согласные звуки — это как раз сонорные. Их пять:
Иногда [Й] не относят к сонорным, но он все равно остается звонким непарным. Посмотрим на таблицу.
По ней видно, что, кроме звонких непарных, имеются и звуки, являющиеся непарными глухими. Большинство из них шипящие; к шипящим не относится только глухой непарный согласный звук [Ц].
В этой статье мы рассматриваем лишь русские звуки речи. В других языках распределение по парам может быть иным. Например, в тибетском языке имеется глухая пара к звонкому [Л].
Пары по твердости-мягкости
Кроме глухости-звонкости, русские согласные образуют пары по твердости-мягкости.
Это значит, что одни из них на слух воспринимаются как более мягкие. Тогда мы обычно как-то обозначаем это на письме: например, пишем мягкий знак или одну из гласных Е, Ё, Ю, Я.
Устная речь первична (любому понятно, что она появилась раньше письменной), поэтому неправильно говорить: «Звук [Н’] в слове КОНЬ мягкий, потому что после него стоит Ь». Напротив, мы пишем Ь, потому что Н’ мягкий.
По твердости-мягкости согласные также составляют пары. Но и в этом случае не все. В русском языке есть непарные мягкие и непарные твердые согласные звуки.
Непарные твердые согласные звуки — это, в основном, шипящие ([Ж], [Ш]) и [Ц]. Они всегда образуются у дальнего нёба.
А вот в предке нашего языка, старославянском, напротив, [Ж] и [Ш] были всегда мягкими и не имели твердой пары. Тогда мягкими не бывали [К], [Г] и [Х]. В настоящее время можно встретить (некогда единственно возможное) произношение с мягким [Ж’] [ДРОЖ’Ж’И] или [ДОЖ’] (дождь), но это теперь необязательно.
Непарные мягкие — это [Й’] и опять же шипящие [Ч’] и [Щ’].
То есть все шипящие являются или всегда твердыми, или всегда мягкими. Буква Ь после них не указывает на мягкость, она выполняет грамматическую функцию (например, даже не зная, что такое «плешь», любой сразу скажет, что это слово женского рода, ведь в мужском роде после шипящих Ь не ставится). Твердые непарные шипящие согласные звуки в слове могут иметь при себе Ь, но это не значит, что они должны смягчиться. Это значит, что перед нами существительное 3 склонения, наречие или глагол.
Непарные мягкие согласные звуки в слове вызывают желание поставить после них Ь, что часто не требуется. Поэтому есть смысл запомнить, что в сочетаниях ЧК, ЧН и т.п. Ь после Ч не нужен.
Звуки «совсем непарные»
В русском языке большинство согласных или парные по обоим признакам, или парные по одному признаку и непарные по другому. Например
, в слове [П’ЭН’] (пень) звук [П’] парный и по глухости-звонкости (П’ – Б’), и по твердости-мягкости (П’ — П), а звук [Н’] парный по твердости-мягкости (Н’ — Н), но непарный по глухости-звонкости.
Однако есть несколько звуков, непарных по обоим признакам. Это звуки [Й’] (непарный звонкий, непарный мягкий), [Ч’] (непарный мягкий, непарный глухой), [Щ’] (непарный мягкий, непарный глухой) и [Ц] (непарный твёрдый, непарный глухой). Такие звуки нередко загадывают в олимпиадах по русскому языку. Например,
«Угадайте звук по характеристике: непарный твёрдый, непарный глухой». Мы уже видим, что это [Ц].
Что мы узнали?
Из статьи про парные и непарные согласные, мы узнали, что в русском языке существуют как парные, так и непарные согласные. Парные согласные различаются по глухости-звонкости и по твердости-мягкости.
Тест по теме
Оценка статьи
Средняя оценка: 4.2
. Всего получено оценок: 209.
Чем отличаются гласные и согласные буквы и звуки? Каким правилам они подчиняются? Как обозначается твердость и мягкость звуков и букв? На все эти вопросы вы получите ответы в представленной статье.
Общие сведения о гласных и согласных буквах
Гласные и согласные буквы представляют собой основу всего русского языка. Ведь при помощи их сочетаний образуются слоги, которые складываются в слова, выражения, предложения, тексты и проч. Именно поэтому данной теме посвящается довольно много часов в средней школе.
и звуки в русском языке
О том, какие гласные и согласные буквы имеются в русском алфавите, человек узнает уже с первого класса. И несмотря на всю кажущуюся простоту данной темы, она считается одной из самых сложных для учеников.
Итак, в русском языке имеется десять гласных букв, а именно: о, и, а, ы, ю, я, э, ё, у, е. Во время их непосредственного произношения можно ощутить, как воздух беспрепятственно проходит через ротовую полость. При этом мы довольно отчетливо слышим свой собственный голос. Следует также отметить, что гласные звуки можно тянуть (а-а-а-а, э-э-э-э-э, и-и-и-и-и, у-у-у-у-у и так далее).
Особенности и букв
Гласные буквы являются основой слога, то есть именно они его организуют. Как правило, в русских словах столько же слогов, сколько и самих гласных. Приведем наглядный пример: у-че-ни-ки — 5 слогов, ре-бя-та — 3 слога, он — 1 слог, о-но — 2 слога и так далее. Бывают даже такие слова, которые состоят лишь из одного гласного звука. Обычно это междометия (А!, О!, У-у-у!) и союзы (и, а и проч.).
Окончаниях, суффиксах и приставках — это очень важные темы в дисциплине «Русский язык». Ведь без знания того, как пишутся такие буквы в том или ином слове, довольно проблематично составить грамотное письмо.
Согласные буквы и звуки в русском языке
Гласные и согласные буквы и звуки значительно различаются. И если первые можно легко тянуть, то последние произносятся максимально коротко (кроме шипящих, так как их можно тянуть).
Следует отметить, что в русском алфавите количество согласных букв — 21, а именно: б, в, г, д, ж, з, й, к, л, м, н, п, р, с, т, ф, х, ц, ч, ш, щ. Звуки, обозначаемые ими, принято делить на глухие и звонкие. Чем они отличаются? Дело в том, что во время произношения звонких согласных человек может слышать не только характерный шум, но и собственный голос (б!, з!, р! и проч.). Что касается глухих, то их никак нельзя произнести громко или, например, прокричать. Они создают только своеобразный шум (ш-ш-ш-ш-ш, с-с-с-с-с и проч.).
Таким образом, практически все подразделяются на две разные категории:
- звонкие — б, в, г, д, ж, з, й, л, м, н, р;
- глухие — к, п, с, т, ф, х, ц, ч, ш.
Мягкость и твердость согласных звуков
Далеко не каждый знает, но гласные и согласные звуки могут быть твердыми и мягкими. Это второй важнейший признак в русском языке (после звонкости и глухости).
Отличительной чертой мягких согласных является то, что во время их произношения язык человека принимает особое положение. Как правило, он сдвигается немного вперед, а вся его средняя часть слегка приподнимается. Что касается то при их произношении язык оттягивается назад. Можете сравнить положение своего речевого органа самостоятельно: [н] — [н’], [т] — [т’]. Следует также отметить, что звонкие и мягкие звуки звучат несколько выше, нежели твердые.
В русском языке практически все согласные имеют пары по признаку мягкости и твердости. Однако есть и такие, у которые их просто нет. К ним относятся твердые — [ж], [ш] и [ц] и мягкие — [й»], [ч»] и [ш»].
Мягкость и твердость гласных звуков
Наверняка мало кто слышал о том, что в русском языке имеются мягкие гласные. Мягкие согласные — это вполне привычные для нас звуки, чего не скажешь о вышеупомянутых. Отчасти это связано с тем, что в средней школе данной теме практически не уделяется время. Ведь и так понятно, при помощи каких гласных согласные становятся мягкими. Однако мы все же решили посвятить вас в эту тему.
Итак, мягкими называют те буквы, которые способны смягчать идущие перед ними согласные. К ним относятся следующие: и, ё, я, е, ю. Что касается таких букв, как а, у, ы, э, о, то они считаются твердыми, так как не смягчают идущие впереди согласные. Чтобы убедиться в этом приведем несколько примеров:
Обозначение мягкости согласных букв при фонетическом разборе слова
Звуки и буквы русского языка изучает фонетика. Наверняка, в средней школе вас не раз просили сделать какого-либо слова. Во время такого анализа следует обязательно указывать на то, является ли отдельно рассматриваемая или нет. Если да, то ее необходимо обозначать следующим образом: [н’], [т’], [д’], [в’], [м’], [п’]. То есть вверху справа рядом с согласной буквой, стоящей перед мягкой гласной, требуется ставить своеобразную черточку. Аналогичным значком помечаются и следующие мягкие звуки — [й»], [ч»] и [ш»].
Все согласные звуки в русском языке разделяются по нескольким признакам, в том числе и по принципу звонкости-глухоты. Эта произносительная характеристика влияет на то, используется ли при выговаривании звука голос или нет. Изучение данной темы очень важно для понимания основных принципов работы фонетической системы, ведь глухие согласные звуки являются очень важной ее частью.
Что такое глухой согласный звук
Глухие согласные звуки производятся только шумом, без участия голоса. При их произнесении голосовые связки полностью расслабляются, гортань не вибрирует.
Парные и непарные глухие согласные
Большинство звуков, которые относятся к этой категории, имеют звонкую пару. Какие это звуки, можно узнать из таблицы “Глухие согласные звуки в русском языке”.
Таким образом, в русском языке есть 11 глухих согласных, имеющих звонкую пару. Но есть и непарные – это такие звуки, как [х], [х’], [ч’] и [щ’].
Они не могут стать звонкими вне зависимости от позиции.
Запомнить все глухие согласные, которые есть в русском языке, помогает специальная мнемоническая фраза: “Степка, хочешь щец? – Фу!”. Но запомнить их парность по твердости-мягкости она не поможет, так как глухие согласные, имеющие пару, представлены в ней только в одной разновидности – или твердой, или мягкой.
Правило оглушения согласных
В русском языке нередки случаи, когда на письме пишется звонкая согласная буква, а в речи она превращается в глухой согласный звук. Это происходит, например, когда звонкая буква оказывается в самом конце слова, как в слове гриб, транскрипция которого будет выглядеть как [грип].
Из-за того, что звонкие согласные на конце оглушаются, часто возникают сложности при воспроизведении таких слов на письме. Однако есть простой способ проверить, какую букву использовать: нужно так изменить слово, чтобы согласная оказалось перед гласной, например, гриб – гриба. Тогда сразу станет понятно, что нужно писать. То же самое касается случаев, когда на конце стоит глухой согласный, а на письме его озвончают “по общему правилу”. Проверить, какая буква пишется, можно тем же способом: крик – крика, лот – лота.
Также могут оглушаться звонкие согласные, располагающиеся в позиции в начале и в середине слова, если после них идет глухой согласный. Это легко понять на примере: будка [б́утка].
Что мы узнали?
Глухие согласные звуки – это такие звуки, при образовании которых не вибрирует гортань, то есть не участвует голос. Состоят они только из шума. У большинства глухих согласных есть звонкая пара, однако есть четыре непарных звука такого типа – это [х], [х’], [ч’] и [щ’]. Из-за правила оглушения согласных при произнесении те согласные, которые на письме являются звонкими, переходят в свою глухую пару. Так происходит, если они оказываются в конце слова, а также когда перед ними стоит другой глухой согласный.
В русском языке 21 согласная буква и 37 согласных звуков:
Согласные звуки бывают твёрдые и мягкие, звонкие и глухие. Мягкость звука обозначается так — [«].
Твёрдые и мягкие согласные звуки
Твёрдый согласный звук получается, если после согласной стоит гласная А, О, У, Ы или Э:
на ло ку мы фэ
Мягкий согласный звук получается, если после согласной стоит гласная Е, Ё, И, Ю или Я:
бе лё ки ню ля
Мягкость согласных звуков так же обозначается с помощью мягкого знака — Ь. Сам мягкий знак звука не обозначает, он пишется после согласной буквы и вместе с ней обозначает один мягкий согласный звук:
рысь
[рыс»], огонь
[огон»], вьюга
[в»й»уга].
Большинству согласных букв соответствует два звука: твёрдый и мягкий, такие согласные называются парными.
Парные согласные по твёрдости — мягкости
:
Но есть согласные буквы, котором соответствует только один из звуков: твёрдый или мягкий, такие согласные называются непарными.
Непарные твёрдые согласные
(всегда твёрдые): Ж [ж], Ш [ш], Ц [ц].
Непарные мягкие согласные
(всегда мягкие): Ч [ч»], Щ [щ»], Й [й»].
В русском языке есть долгий звонкий мягкий звук [ж»]. Он встречается в небольшом количестве слов и получается только при произнесении сочетаний букв жж, зж, жд
: вожжи, дребезжать, дождь
.
Звонкие и глухие согласные звуки
Согласные звуки можно разделить на глухие и звонкие.
Глухими согласными называются такие звуки, при произношении которых не используется голос, они состоят только из шума, например: звуки [с], [ш], [ч»].
Звонкими согласными называются такие звуки, при произношении которых используется голос, то есть они состоят из голоса и шума, например: звуки [р], [ж], [д].
Некоторые звуки составляют пару: звонкий — глухой, такие звуки называются парными.
Парные согласные по глухости — звонкости
:
Непарные звонкие согласные
: Й, Л, М, Н, Р.
Непарные глухие согласные
: Х, Ц, Ч, Щ.
Шипящие и свистящие согласные звуки
Звуки [ж], [ш], [ч»], [щ»] называются шипящими согласными звуками. Звуки [ж] и [ш] — это непарные твёрдые шипящие согласные звуки:
жук
[жук], шут
[шут]
Звуки [ч»] и [щ»] — это непарные мягкие шипящие согласные звуки:
чиж
[ч»иж], щит
[щ»ит]
Звуки [з], [з»] [с], [с»], [ц] называются свистящими согласными звуками.
Буква и звук Й
Буква Й (и краткое) обозначает звук [й»]: рай [рай»]. Буква Й пишется:
- В начале слов: йод, йогурт.
- В середине слов, перед согласными буквами: лайка, майка, кофейник.
- В конце слов: рай, май, твой.
Звук [й»] встречается чаще буквы Й, так как он появляется в словах где нет буквы Й, но есть гласные Я, Е, Ю и Ё. Рассмотрим в каких случаях звук [й»] встречается в словах, не содержащих букву Й:
- гласные Я, Е, Ю и Ё стоят в начале слова: яма
[й»ама
] - гласные Я, Е, Ю и Ё стоят после гласных: дует
[дуй»эт
] - гласные Я, Е, Ю и Ё стоят после разделительного твёрдого знака (Ъ): въезд
[вй»эзд
] - гласные Я, Е, Ю и Ё стоят после разделительного мягкого знака (Ь): льёт
[л»й»от
] - гласная И стоит после разделительного мягкого знака (Ь): ульи
[ул»й»и
]
1 Организационный | Организация рабочего места | Приветствует детей: У нас на парте всё в порядке: Альбомы, книги и тетрадки. Готовы мы начать урок, Звени скорей для нас звонок! Проверим готовность к уроку Называет учебные принадлежности к уроку | Проверяют готовность к уроку. Карточки пока не трогают | Личностные: Проявляют эмоциональную отзывчивость на слова учителя. Правильно идентифицируют себя с позиций школьника. | 2. Актуализация знаний | Здесь зашифрованы слова. Подумайте, как прочитать. Попробуйте прочитать и вы узнаете, о чем пойдет речь на уроке. (слайд на экране) (Ключевые слова: парные, непарные) О каких звуках можно сказать: парные – непарные? Что вы знаете об этом? Как можно объединить согласные по парности? (звонкость-глухость, мягкость-твердость). Какой из признаков – постоянный? От чего зависит мягкость-твердость согласных? Назовите всегда твердые согласные, всегда мягкие согласные. | Организует мыслительные операции. Формулирует задания. Активизирует знания учащихся. Создает проблемную ситуацию. Нацеливает на воспроизведение известных знаний | Высказываются. Вспоминают известные сведения Делают вывод о парности согласных. Выбирают нужные знания на данном уроке. | Предметные (логические): Осуществляют поиск необходимой информации. Регулятивные: действуют с учетом выделенных ориентиров. | 3. Организация познавательной деятельности | Слайд на экране. Посмотрите на буквы, они подскажут, с какими парами согласных будем работать на уроке. Ь Я Е Ё Ю И О чем говорят эти буквы? Обсуди с соседом по парте, что объединяет эти буквы? Сделайте вывод. Какая буква отличается от других? Что вы знаете о ней? | После высказывания детей более полно проговаривает тему урока, определяет цель урока. Предлагает выполнить задание в парах. | Целеполагание : формулируют тему и цель урока. В парах обсуждают друг с другом и проговаривают сходство и различие. Делают вывод. | Предметные: осуществлять поиск информации, подведение под понятие, выделение существенных признаков. Регулятивные: принимать и сохранять учебную задачу. Коммуникативные: формулировать собственное мнение. Строить понятные высказывания, слушать собеседника | 4. Работа по теме урока | 1.Работа по учебнику стр. 83, упр. 5. Произносите каждую пару звуков. Какие различия при звучании вы заметили? Определить, какие твердые, а какие мягкие звуки. Одинаково ли они обозначаются на письме. Найти и назвать звуки, не имеющие пары. Определить их по твердости и мягкости. 2. Прослушиваем электронное приложение к учебнику. Урок 14 «Твердые и мягкие согласные звуки» 3. Стр. 83, упр.5. Для чего в таблице даны гласные буквы? Какой гласной там нет? Где бы вы ее расположили – справа или слева? Почему? | Организация восприятия и осмысления новой информации: усвоение исходных знаний | Произносят каждую пару звуков. Определяют, одинаково ли они звучат и обозначаются на письме. Формулируют выводы. | Предметные: воспроизводить информацию по памяти, необходимую для решения учебной задачи, приводить примеры, в качестве доказательства выдвигаемых положений и выводов. | 5.Физминутка | Игра «Звуковая мозаика».(слова с парными согласными и без них) Прыгаем, если слышим на конце слова парный согласный. Приседаем, если этой орфограммы нет. Слова: корова, снег, парус, глаза, ландыш, хлеб, лопата, мед, пилот, береза | Проводит физминутку | Выполняют упражнения | Регулятивные: осуществляют пошаговый контроль своих действий, ориентируясь на показ движений учителем. | 6. Первичная проверка понимания | 1.Рабочая тетрадь, с.42 упр.1 Прочитайте слова. Какими буквами различается каждая пара слов? (гласными) А какими звуками различаются? Почему? Произнесите эти звуки. Раскрасьте схемы нужными цветами. Проверка: показать синюю карточку, если звук твердый и зеленую, если звук мягкий. 2. Индивидуальная работа (3 ученика) работают по карточкам. Задание: подбери и запиши слова, в которых есть такие звуки: 1 ученик- [c], [c,], [м], [м,], 2 ученик — [н], [н,], [л], [л,], 3 ученик — [р], [р,], [т], [т,]. 3.Упр. 6, стр.84 Прочитайте стихотворение. Определите главную мысль. Спишите первое предложение, подчеркните буквы, которые указывают на мягкость предшествующего согласного. Какие это будут буквы? Взаимопроверка. 4.Словарная работа — Кто не хочет войны? (ребята). — Что обозначает это слово? (слайд) — Подберите близкие по значению слова. — Прочитай слово с ударением. — Сколько в нем слогов? — Найдите «опасные» места в слове, объясните. — Запишите слово, разделив для переноса. | Организует усвоение способов деятельности путем воспроизведения информации и упражняет в ее применении | Читают слова, произносят звуки, раскрашивают схемы. Читают. Отвечают на вопросы Работа в парах, обсуждение выполненного задания. Работают по карточкам. Один ученик у доски моделирует слово с помощью схемы Запись слова Самостоятельная работа в тетради | Предметные: из собственного опыта осуществлять поиск необходимой информации Коммуникативные: обмениваются мнениями, слушают друг друга, строят понятные речевые высказывания. Регулятивные: действуют с учетом учителем ориентиров, ориентируются в учебнике и тетради |
Аудиовизуальный опыт усиливает мультисенсорные сборки в зрительной коре головного мозга взрослых мышей
Феллеман Д. Дж. И Ван Эссен Д. С. Распределенная иерархическая обработка в коре головного мозга приматов. Cereb. Кортекс Н. Ю. Н. 1991 1 , 1–47 (1991).
CAS
Google Scholar
Джонс, Э. Дж. И Пауэлл, Т. П. Анатомическое исследование сходящихся сенсорных путей в коре головного мозга обезьяны. Brain J. Neurol. 93 , 793–820 (1970).
CAS
Статья
Google Scholar
Фишман М. К. и Майкл П. Интеграция слуховой информации в зрительную кору головного мозга кошки. Vis. Res. 13 , 1415–1419 (1973).
CAS
PubMed
Статья
Google Scholar
Ибрагим, Л. А. и др. Усиление кросс-модальности зрительной корковой обработки посредством ингибирования и растормаживания, опосредованных слоем 1. Neuron 89 , 1031–1045 (2016).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Iurilli, G. et al. Управляемое звуком синаптическое торможение в первичной зрительной коре. Нейрон 73 , 814–828 (2012).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Кайзер, К., Петков, К. И., Аугат, М., Логотетис, Н. К. Интеграция прикосновения и звука в слуховой коре. Нейрон 48 , 373–384 (2005).
CAS
PubMed
Статья
Google Scholar
Meijer, G. T., Montijn, J. S., Pennartz, C. M. A. & Lansink, C. S. Аудиовизуальная модуляция в первичной зрительной коре мышей зависит от кросс-модальной конфигурации стимула и конгруэнтности. J. Neurosci. J. Soc.Neurosci. 37 , 8783–8796 (2017).
CAS
Статья
Google Scholar
Моррелл Ф. Вид акустического пространства визуальной системой. Nature 238 , 44–46 (1972).
ADS
CAS
PubMed
Статья
Google Scholar
Piché, M. et al. Слуховые ответы в зрительной коре неонатально энуклеированных крыс. Neuroscience 145 , 1144–1156 (2007).
PubMed
Статья
CAS
Google Scholar
Спинелли, Д. Н., Старр, А. и Барретт, Т. В. Слуховая специфичность в записях единиц зрительной коры головного мозга кошки. Exp. Neurol. 22 , 75–84 (1968).
CAS
PubMed
Статья
Google Scholar
Чандрасекаран, К.Вычислительные принципы и модели мультисенсорной интеграции. Curr. Opin. Neurobiol. 43 , 25–34 (2017).
CAS
PubMed
Статья
Google Scholar
Урсино, М., Куппини, К. и Магоссо, Э. Нейрокомпьютерные подходы к моделированию мультисенсорной интеграции в головном мозге: обзор. Нейронная сеть. J. Int. Neural Netw. Soc. 60 , 141–165 (2014).
МАТЕМАТИКА
Статья
Google Scholar
Atilgan, H. et al. Интеграция визуальной информации в слуховой коре способствует анализу слуховых сцен за счет мультисенсорной привязки. Neuron 97 , 640–655.e4 (2018).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Mezzera, C. & López-Bendito, G. Кросс-модальная пластичность в моделях животных с сенсорной депривацией: с точки зрения таламокортикального развития. Дж.Chem. Нейроанат. 75 , 32–40 (2016).
PubMed
Статья
Google Scholar
Yu, L., Xu, J., Rowland, B. A. и Stein, B. E. Мультисенсорная пластичность в нейронах верхнего бугорка опосредуется ассоциативной корой. Cereb. Кортекс Н. Ю. Н. 1991 26 , 1130–1137 (2016).
Google Scholar
Доррн, А.Л., Юань, К., Баркер, А. Дж., Шрейнер, К. Э. и Фремке, Р. К. Сенсорный опыт развития уравновешивает корковое возбуждение и торможение. Природа 465 , 932–936 (2010).
ADS
CAS
PubMed
PubMed Central
Статья
Google Scholar
Ниелл, К. М. и Страйкер, М. П. Высокоселективные рецептивные поля в зрительной коре головного мозга мышей. J. Neurosci. J. Soc. Neurosci. 28 , 7520–7536 (2008).
CAS
Статья
Google Scholar
Келлер, Г. Б., Бонхёффер, Т. и Хюбенер, М. Сигналы сенсомоторного несоответствия в первичной зрительной коре играющей мыши. Нейрон 74 , 809–815 (2012).
CAS
PubMed
Статья
Google Scholar
Akemann, W. et al. Двухфотонная визуализация напряжения с использованием генетически закодированного индикатора напряжения. Sci. Отчет 3 , 2231 (2013).
PubMed
PubMed Central
Статья
Google Scholar
Эмпсон, Р. М. и др. Проверка данных об оптическом напряжении с помощью генетически закодированного индикатора напряжения VSFP-Butterfly от пирамидных нейронов кортикального слоя 2/3 в срезах мозга мышей. Physiol. Репу . 3 , e12468 (2015). [Epub, 29 июля 2015 г.]
PubMed
PubMed Central
Статья
Google Scholar
Song, C., Barnes, S. & Knöpfel, T. Визуализация напряжения коры мозга млекопитающих с использованием генетически закодированных индикаторов напряжения: обзор, посвященный профессору Амираму Гринвальду. Нейрофотоника 4 , 031214 (2017).
PubMed
PubMed Central
Статья
Google Scholar
Barnes, S.J. et al. Специфическая для подсети гомеостатическая пластичность в зрительной коре головного мозга мышей in vivo. Нейрон 86 , 1290–1303 (2015).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Ko, H. et al. Функциональная специфика локальных синаптических связей в неокортикальных сетях. Природа 473 , 87–91 (2011).
ADS
CAS
PubMed
PubMed Central
Статья
Google Scholar
Миллер, Дж. К., Айзенштат, И., Каррильо-Рид, Л.& Юсте, Р. Визуальные стимулы задействуют внутренне сгенерированные корковые ансамбли. Proc. Natl Acad. Sci. США 111 , E4053 – E4061 (2014).
CAS
PubMed
Статья
Google Scholar
Йошимура, Ю., Данцкер, Дж. Л. М., Каллауэй, Э. М. Возбуждающие корковые нейроны образуют мелкомасштабные функциональные сети. Природа 433 , 868–873 (2005).
ADS
CAS
PubMed
Статья
Google Scholar
Ch’ng, Y.H. & Reid, R.C. Клеточная визуализация зрительной коры головного мозга выявляет пространственную и функциональную организацию спонтанной активности. Фронт. Интегр. Neurosci . 4 , pii: 20. https://doi.org/10.3389/fnint.2010.00020 (2010).
Hofer, S. B. et al. Дифференциальная связность и динамика ответа возбуждающих и тормозных нейронов зрительной коры. Nat. Neurosci. 14 , 1045–1052 (2011).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Komiyama, T. et al. Связанная с обучением мелкомасштабная специфичность, отображаемая в цепях моторной коры у ведущих мышей. Природа 464 , 1182–1186 (2010).
ADS
CAS
PubMed
Статья
Google Scholar
Биненшток, Э. Л., Купер, Л. Н. и Манро, П. В. Теория развития селективности нейронов: специфичность ориентации и бинокулярное взаимодействие в зрительной коре головного мозга. J. Neurosci.J. Soc. Neurosci. 2 , 32–48 (1982).
CAS
Статья
Google Scholar
Суини, Ю., Барнс, С. и Клопат, К. Разнообразные гомеостатические реакции на визуальную депривацию путем выявления повторяющихся подсетей. bioRxiv 312926. https://doi.org/10.1101/312926 (2018)
Яо, Х. и Дэн, Ю. Пластичность, зависящая от времени стимула в корковой обработке ориентации. Neuron 32 , 315–323 (2001).
CAS
PubMed
Статья
Google Scholar
Сюй, С., Цзян, В., Пу, М.-М. & Дэн, Ю. Напоминание активности в визуальном кортикальном ансамбле. Nat. Neurosci. 15 , 449–455 (2012). S1-2.
CAS
PubMed
PubMed Central
Статья
Google Scholar
Яо, Х., Ши, Л., Хань, Ф., Гао, Х. и Дан, Ю. Быстрое обучение кортикальному кодированию визуальных сцен. Nat. Neurosci. 10 , 772–778 (2007).
CAS
PubMed
Статья
Google Scholar
Маффеи, Л., Фиорентини, А. и Бисти, С. Нейронный коррелят перцептивной адаптации к решеткам. Наука 182 , 1036–1038 (1973).
ADS
CAS
PubMed
Статья
Google Scholar
Eysel, U. T., Eyding, D. & Schweigart, G. Повторяющаяся оптическая стимуляция вызывает быстрые изменения рецептивного поля в зрелой зрительной коре. Нейроотчет 9 , 949–954 (1998).
CAS
PubMed
Статья
Google Scholar
Хан, Ф., Капорале, Н. и Дэн, Ю. Отражение недавнего визуального опыта в спонтанных корковых волнах. Нейрон 60 , 321–327 (2008).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Андерманн, М. Л., Керлин, А. М. и Рид, Р. С. Хроническая клеточная визуализация зрительной коры мышей во время оперантного поведения и пассивного просмотра. Фронт. Клетка. Neurosci. 4 , 3 (2010).
PubMed
PubMed Central
Google Scholar
Кук, С. Ф. и Беар, М. Ф. Пластичность стимулов-избирательного ответа в зрительной коре: анализ для оценки патофизиологии и лечения когнитивных нарушений, связанных с психическими расстройствами. Biol. Психиатрия 71 , 487–495 (2012).
PubMed
Статья
Google Scholar
Кук, С. Ф. и Беар, М. Ф. Память визуального распознавания: взгляд из V1. Curr. Opin. Neurobiol. 35 , 57–65 (2015).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Кук, С. Ф., Коморовски, Р.W., Kaplan, E. S., Gavornik, J. P. и Bear, M. F. Зрительная память распознавания, проявляющаяся как долгосрочное привыкание, требует синаптической пластичности в V1. Nat. Neurosci. 18 , 262–271 (2015).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Гаворник, Дж. П. и Медведь, М. Ф. Распознавание и предсказание выученной пространственно-временной последовательности в первичной зрительной коре. Nat. Neurosci. 17 , 732–737 (2014).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Бенуччи А., Салим А. Б. и Карандини М. Адаптация поддерживает гомеостаз населения в первичной зрительной коре. Nat. Neurosci. 16 , 724–729 (2013).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Keller, A.J. et al. Релевантность стимула модулирует адаптацию контраста в зрительной коре. eLife 6 , pii: e21589. https://doi.org/10.7554/eLife.21589 (2017).
Като, Х. К., Жиллет, С. Н. и Исааксон, Дж. С. Гибкие сенсорные представления в слуховой коре, обусловленные поведенческой релевантностью. Neuron 88 , 1027–1039 (2015).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Натан, Р. Г. и др. Дополнительный контроль сенсорной адаптации двумя типами корковых интернейронов. eLife 4 , pii: e09868. https://doi.org/10.7554/eLife.09868 (2015).
Cossell, L. et al. Функциональная организация возбуждающей синаптической силы в первичной зрительной коре. Природа 518 , 399–403 (2015).
ADS
CAS
PubMed
PubMed Central
Статья
Google Scholar
Хан, А.Г. и др. Отчетливые вызванные обучением изменения в селективности стимулов и взаимодействия классов ГАМКергических интернейронов в зрительной коре. Nat. Neurosci. 21 , 851–859 (2018).
CAS
PubMed
Статья
Google Scholar
Сугихара, Х., Чен, Н. и Сур, М. Клеточно-специфическая модуляция пластичности и коркового состояния посредством холинергических входов в зрительную кору. J. Physiol.Париж 110 , 37–43 (2016).
PubMed
PubMed Central
Статья
Google Scholar
Кайко-Гаджик, Н. А. и Суини, Ю. Углубляясь в кросс-модальную интеграцию. J. Neurosci. J. Soc. Neurosci. 38 , 6442–6444 (2018).
CAS
Статья
Google Scholar
Кампа, Б. М., Рот, М. М., Гёбель, В.И Хельмхен, Ф. Представление визуальных сцен локальными популяциями нейронов в слое 2/3 зрительной коры головного мозга мыши. Фронт. Нейронные схемы 5 , 18 (2011).
PubMed
PubMed Central
Статья
Google Scholar
Райхе, Р. В. и Сур, М. Пространственные корреляции в естественных сценах модулируют надежность реакции в зрительной коре головного мозга мыши. J. Neurosci. J. Soc. Neurosci. 35 , 14661–14680 (2015).
CAS
Статья
Google Scholar
Харрис, К. Д. и миссис-Флогель, Т. Д. Кортикальные связи и сенсорное кодирование. Природа 503 , 51–58 (2013).
ADS
CAS
PubMed
Статья
Google Scholar
Джи Д. и Уилсон М. А. Скоординированное воспроизведение памяти в зрительной коре и гиппокампе во время сна. Nat.Neurosci. 10 , 100–107 (2007).
CAS
PubMed
Статья
Google Scholar
Канг, Дж. И., Хуппе-Гург, Ф. и Вошер, Э. Повышение функции и пластичности зрительной коры головного мозга с помощью ацетилхолина для улучшения зрительного восприятия. Фронт. Syst. Neurosci . 8 , 172. https://doi.org/10.3389/fnsys.2014.00172 (2014).
Пфеффер, К. К., Сюэ, М., Хе, М., Хуанг, З.J. & Scanziani, M. Ингибирование торможения в зрительной коре: логика связей между молекулярно различными интернейронами. Nat. Neurosci. 16 , 1068–1076 (2013).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Gentet, L.J. Функциональное разнообразие супрагранулярных ГАМКергических нейронов в бочкообразной коре. Фронт. Нейронные схемы 6 , 52 (2012).
PubMed
PubMed Central
Статья
CAS
Google Scholar
Hensch, T. K. et al. Локальный контроль цепи ГАМК зависимой от опыта пластичности в развитии зрительной коры. Наука 282 , 1504–1508 (1998).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Андерманн, М. Л., Керлин, А.М., Румис, Д. К., Гликфельд, Л. Л. и Рид, Р. С. Функциональная специализация верхних зрительных областей коры мозга у мышей. Neuron 72 , 1025–1039 (2011).
CAS
PubMed
Статья
Google Scholar
Аяз А., Салим А. Б., Шельвинк М. Л. и Карандини М. Локомоция контролирует пространственную интеграцию в зрительной коре головного мозга мыши. Curr. Биол. CB 23 , 890–894 (2013).
CAS
PubMed
Статья
Google Scholar
Fiser, A. et al. Зависящие от опыта пространственные ожидания в зрительной коре головного мозга мыши. Nat. Neurosci. 19 , 1658–1664 (2016).
CAS
PubMed
Статья
Google Scholar
Ниелл, К. М. и Страйкер, М. П. Модуляция зрительных ответов поведенческим состоянием в зрительной коре головного мозга мыши. Нейрон 65 , 472–479 (2010).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Pakan, J. M. et al. Модуляция ингибирования поведенческим состоянием зависит от контекста и зависит от типа клеток в зрительной коре головного мозга мыши. eLife 5 , pii: e14985. https://doi.org/10.7554/eLife.14985 (2016).
Roth, M. M. et al. Ядра таламуса передают разнообразную контекстную информацию в слой 1 зрительной коры. Nat. Neurosci. 19 , 299–307 (2016).
CAS
PubMed
Статья
Google Scholar
Салим, А. Б., Аяз, А., Джеффри, К. Дж., Харрис, К. Д. и Карандини, М. Интеграция зрительного движения и передвижения в зрительной коре головного мозга мыши. Nat. Neurosci. 16 , 1864–1869 (2013).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Tohmi, M., Meguro, R., Tsukano, H., Hishida, R. & Shibuki, K. Экстрагенный зрительный путь генерирует отличные свойства ответа в высших зрительных областях мышей. Curr. Биол. CB 24 , 587–597 (2014).
CAS
PubMed
Статья
Google Scholar
Yoshitake, K. et al. Сдвиг визуальной карты на основе сигналов, управляемых усами в зрительной коре головного мозга молодых мышей. Cell Rep. 5 , 1365–1374 (2013).
CAS
PubMed
Статья
Google Scholar
Zmarz, P. & Keller, G.Б. Несовпадение рецептивных полей в зрительной коре головного мозга мыши. Neuron 92 , 766–772 (2016).
CAS
PubMed
Статья
Google Scholar
Holtmaat, A. et al. Долгосрочная визуализация с высоким разрешением в неокортексе мыши через хроническое черепное окно. Nat. Protoc. 4 , 1128–1144 (2009).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Lissek, T. et al. Общие анестезиологические условия вызывают сетевую синхронизацию и нарушают обработку сенсорной информации в коре головного мозга. Фронт. Клетка. Neurosci . 10 , 64 (2016).
Barnes, S.J. et al. Индуцированное депривацией гомеостатическое масштабирование позвоночника in vivo локализовано в дендритных ветвях, которые недавно подверглись потере позвоночника. Neuron 96 , 871–882.e5 (2017).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Real, R. et al. Моделирование динамики нейронов человека и синдрома Дауна in vivo. Наука . https://doi.org/10.1126/science.aau1810 (2018)
ADS
PubMed
PubMed Central
Статья
CAS
Google Scholar
Sammons, R.P., Clopath, C. & Barnes, S.J. Зависимая от размера динамика аксонального бутона после визуальной депривации in vivo. Cell Rep. 22 , 576–584 (2018).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Дуббс, А., Гевара, Дж. И Юсте, Р. Моко: коррекция быстрого движения для визуализации кальция. Фронт. Нейроинформатика 10 , 6 (2016).
Carandini, M. et al. Визуализация бодрствующей зрительной коры с помощью генетически закодированного индикатора напряжения. J. Neurosci. J. Soc. Neurosci. 35 , 53–63 (2015).
CAS
Статья
Google Scholar
Хеннекен Г., Фогельс Т.П. и Герстнер, В. Оптимальное управление динамикой переходных процессов в сбалансированных сетях поддерживает генерацию сложных движений. Нейрон 82 , 1394–1406 (2014).
CAS
PubMed
PubMed Central
Статья
Google Scholar
Раджан, К., Эбботт, Л. Ф. и Сомполинский, Х. Стимул-зависимое подавление хаоса в рекуррентных нейронных сетях. Phys. Rev. E Stat. Нелин. Soft Matter Phys. 82 , 011903 (2010).
ADS
PubMed
Статья
CAS
Google Scholar
Фогельс, Т. П., Спрекелер, Х., Зенке, Ф., Клопат, К. и Герстнер, В. Тормозная пластичность уравновешивает возбуждение и торможение в сенсорных путях и сетях памяти. Наука 334 , 1569–1573 (2011).
ADS
CAS
PubMed
Статья
Google Scholar
Ricciardi, L. M. & Sacerdote, L. Процесс Орнштейна-Уленбека как модель нейрональной активности. I. Среднее и дисперсионное время обжига. Biol. Киберн. 35 , 1–9 (1979).
CAS
PubMed
МАТЕМАТИКА
Статья
Google Scholar
Хрящи гортани — парные — непарные
Гортань (голосовой ящик) — орган, расположенный в передней части шеи. Он является компонентом дыхательных путей и выполняет несколько важных функций, включая фонацию, кашлевой рефлекс и защиту нижних дыхательных путей.
В гортани девять хрящей; три непарных и шесть парных. Они образуют каркас гортани, обеспечивающий жесткость и устойчивость. В этой статье мы рассмотрим анатомию хрящей гортани.
Непарные хрящи
Три непарных хряща — это надгортанника, , щитовидной железы, и перстневидный хрящ, .
Щитовидный хрящ
Щитовидный хрящ — это большая, выступающая структура, хорошо заметная у взрослых мужчин.Он состоит из двух пластин (пластинок), которые соединяются спереди, образуя выступ гортани (кадык).
Задняя граница каждого листа выступает вверх и вниз, образуя верхних и нижних рогов (также известных как корню). Верхние рога соединяются с подъязычной костью через боковую щитовидно-подъязычную связку, а нижние рога сочленяются с перстневидным хрящом.
Перстневидный хрящ
Перстневидный хрящ представляет собой полное кольцо из гиалинового хряща , состоящее из широкого листа сзади и гораздо более узкой дуги спереди (по форме напоминающей перстень с печаткой).
Хрящ полностью окружает дыхательные пути, отмечая нижнюю границу гортани на уровне C6. Он сочленяется с парными черпаловидными хрящами сзади, а также обеспечивает прикрепление нижних рогов щитовидного хряща.
Перстневидный хрящ — это единственный полный хрящевой круг в гортани или трахее. Это имеет клиническое значение во время экстренной интубации, так как давление на перстневидный хрящ может перекрыть пищевод и, таким образом, предотвратить регургитацию желудочного содержимого (известное как давление перстневидного хряща или маневр Селлика).
Надгортанник
Надгортанник представляет собой пластину в форме листа из эластичного хряща , которая отмечает вход в гортань. Его «стебель» прикреплен к задней части переднего отдела щитовидного хряща. Во время глотания надгортанник уплощается и перемещается кзади, чтобы закрыть гортань и предотвратить аспирацию.
Рис. 1.0 — Структура (а) щитовидного хряща и (б) перстневидного хряща. [/ caption]
Парные хрящи
Есть три парных хряща — черпаловидный, роговой и клиновидный.Они расположены в гортани с двух сторон.
Аритеноидные хрящи
Черпаловидные хрящи представляют собой структуры пирамидальной формы, которые располагаются на перстневидном хряще . Они состоят из вершины, основания, трех сторон и двух отростков и служат точкой крепления для различных ключевых структур в гортани:
- Apex — сочленяется с роговым хрящом.
- Основание — сочленяется с верхней границей перстневидного хряща.
- Голосовой отросток — обеспечивает прикрепление голосовой связки.
- Мышечный отросток — обеспечивает прикрепление задних и боковых перстневидных мышц.
Роговые хрящи
Роговые хрящи — это второстепенные хрящевые образования. Они сочленяются с вершинами черпаловидных хрящей.
Клиновидные хрящи
Клиновидные хрящи расположены в пределах надгортанника.У них нет прямого прикрепления, но они укрепляют складки.
Рис. 1.1 — Основные хрящи в скелете гортани. [/ caption]
Стимуляция блуждающего нерва в сочетании с тонами для лечения шума в ушах: проспективное рандомизированное двойное слепое контролируемое пилотное исследование на людях
Дизайн исследования
Исследование представляло собой двойное слепое рандомизированное контролируемое исследование с участием 30 участников. с хроническим нейросенсорным тиннитусом.Участникам обеих групп имплантировали устройство VNS и рандомизировали в парную группу VNS (n = 16) или контрольную (n = 14) группу (рис. 3). Участники проводили лечение дома примерно 2,5 часа в день, 7 дней в неделю, в течение 6 недель. В течение первых 6 недель группа VNS получала VNS в паре с тональными сигналами (обсуждается ниже). Контрольная группа также получала VNS, но не имела пары по тонам. Через 6 недель участники парной группы VNS продолжали получать парные VNS, в то время как участники контрольной группы переходили, чтобы получать парные VNS.Обе группы наблюдались до года.
Рисунок 3
Дизайн исследования. Участникам имплантировали устройство и рандомизировали в парную группу VNS (n = 16) или контрольную (n = 14) группу. Участники проводили лечение дома примерно 2,5 часа в день, 7 дней в неделю. В течение первых 6 недель контрольная группа с имплантированной имплантацией также получала VNS, но не была парной по тонам. После 6 недель слепого контролируемого лечения участники контрольной группы перешли на получение парных VNS.Через 6 недель участники обеих групп получали парную стимуляцию VNS и оценивались в течение одного года.
участников были зарегистрированы в 4 центрах в США (дополнительная таблица 1). Испытание проводилось в рамках утвержденного FDA исключения для исследовательских устройств (IDE, # G130140) и зарегистрировано на сайте Clinicaltrials.gov (NCT01962558) 9 -го октября 2013 г. Информированное согласие было получено в соответствии с требованиями, изложенными в США.Управление по санитарному надзору за качеством пищевых продуктов и медикаментов, Свод федеральных нормативных актов, раздел 21.
Что касается безопасности, выборка из 30 участников позволила достаточно мощности для выявления случаев редких событий, связанных с безопасностью и устройством. Выборка из 30 участников дала 95% вероятность того, что исследование выявит по крайней мере одно проявление всех событий или осложнений, которые происходят у участников, с частотой 9,5% или выше. Кроме того, имплантация и последующее наблюдение 30 участников в течение 6 недель дадут 1260 дней воздействия.В этом случае порог обнаружения снижается до очень маловероятного события, которое происходит всего за 0,2375% дней. Другими словами, если вероятность события составляет 0,002375 или 1 событие за 422 дня воздействия, у нас есть 95% вероятность наблюдения этого события в выборке из 30 участников с 1260 днями воздействия. По мере того, как участники продолжают долгосрочную часть исследования, способность обнаруживать редкие события возрастает по мере увеличения воздействия.
Участники и зачисление
Шестьдесят два участника были отобраны на всех участках исследования.Из 62 участников 32 не продолжили лечение из-за (1) решения участников, как правило, из-за беспокойства участников или члена семьи по поводу хирургического вмешательства или временных затрат на исследование (15), прекращения лечения участком после того, как был достигнут предел имплантата исследования ( 5), несоблюдение текущего уровня шума в ушах (4), противопоказания к приему лекарств (3), другие медицинские проблемы (3) и чрезмерная потеря слуха (2). Тридцать участников были набраны и имплантированы, включая 25 мужчин и 5 женщин.
Критерии включения и исключения
Ключевые критерии включения включали людей с сенсоневральным тиннитусом, которые (a) были в возрасте от 22 до 65 лет (b) имели в основном тональное качество звука в ушах (c) имели односторонний или двусторонний шум в ушах ( г) испытал тиннитус в течение как минимум одного года (д) участвовал как минимум в одной программе терапии тиннитуса и сочел ее бесполезной.Ключевые критерии исключения включали (а) острый или перемежающийся шум в ушах (б) болезнь Меньера, ретрокохлеарную болезнь или признаки активного заболевания среднего уха (в) любое активное имплантированное устройство, такое как кардиостимулятор или другой нейростимулятор, или любое другое исследуемое устройство или лекарство. (d) Перечень депрессии Бека (BDI) 30 или выше (e) Любой препарат, который, как известно, имитирует, увеличивает или уменьшает высвобождение или удаление диффузного нейромодулятора, например норадреналин, дофамин, серотонин, бензодиазепины, ацетилхолин, психоактивные препараты или лекарства. известно, что вызывает или усиливает шум в ушах.
Рандомизация и ослепление
Контрактная исследовательская организация (CRO), выполняющая анализ данных, предоставила рандомизацию для каждого участника на сайт посредством телефонного звонка с неслепым персоналом сайта, который выполнил программирование устройства и настроил устройство на правильные групповые настройки устройства. . Участник инициировал каждый сеанс терапии, но не знал о распределении по группе (была кнопка «Пуск», но участнику не была показана информация о конкретных настройках).Это позволяло поддерживать состояние слепоты, поскольку участники часто не могли воспринимать VNS или только спорадически воспринимали VNS либо через покалывание в шее, либо через охриплость голоса. Аудиологу, проводившему оценку конкретного участника, не было разрешено управлять настройками устройства для этого участника, чтобы оставаться в курсе настроек этого участника. Участники были рандомизированы с помощью SAS (PROC PLAN) и стратифицированы по участкам. У каждого сайта была своя собственная последовательность рандомизации с начальным блоком из двух, за которым следуют блоки из четырех.Для каждого участника сайт вызывал CRO для получения назначения лечения, последовательно взятого из списка рандомизации для этого сайта.
Параметры имплантации и стимуляции VNS
Имплантация устройства обычно выполнялась отоларингологом под общим наркозом. Однако одному участнику по его просьбе была имплантирована местная анестезия. Подробности имплантации устройства были описаны ранее , 28, . После хирургической имплантации и примерно одной недели восстановления участники были рандомизированы либо в парную группу VNS (n = 16), либо в группу активной контрольной терапии (n = 14).
Участники были госпитализированы в хирургический центр / больницу утром в день имплантации устройства и были выписаны в течение 24 часов. Имплантация заключалась в размещении электродов стимуляции на левом блуждающем нерве в сонной артерии. Затем соединитель отведения вводили подкожно в карман, созданный в левой вспомогательной или грудной области, где он был прикреплен к имплантируемому генератору импульсов.
Устройство состояло из имплантируемого генератора импульсов (модель 1000 Serenity®), имплантируемого электрода и электрода (модель 3000) и системы внешнего контроллера (рис.4). Система внешнего контроллера включала портативный компьютер (Dell Inspiron) с высококачественными циркулярными наушниками (Sennheiser, HD280-PRO), на котором запущено программное обеспечение для программирования Tinnitus Application Programming (TAPS Model 4000) и внешний контроллер. Внешний контроллер (модель 2000, подключенный к портативному компьютеру через USB) имел беспроводную связь со стимулятором IPG. Программное обеспечение позволило аудиологу запрограммировать параметры стимуляции (амплитуда (мА), частота (Гц), ширина импульса (мкс), продолжительность (мс)), просмотреть записанную историю программирования участников, а также проверить импеданс отведения и состояние батареи.Программное обеспечение также фиксировало историю программирования участников.
Рисунок 4
Настройка с использованием системы Serenity®, которая сочетает стимуляцию блуждающего нерва (VNS) с тональными сигналами. На вставке показан вывод электрода, обернутый вокруг шейного блуждающего нерва. Устройство представляет собой генератор импульсов, который имплантируется под грудную стенку. Вывод имплантата подключается к генератору импульсов. Изображение любезно предоставлено MicroTransponder , Inc .
Участники начали терапию примерно через неделю восстановления после операции.Стимуляция производилась на левый блуждающий нерв, так как это наиболее распространенная практика при ВНС при эпилепсии и депрессии. Однако, поскольку вышележащие мишени двусторонние, стимуляция, вероятно, влияет на обе стороны полушарий головного мозга. Каждая стимуляция VNS состояла из пятнадцати импульсов постоянного тока по 0,8 мА со сбалансированным зарядом (ширина импульса 100 мкс при 30 Гц). Длительность серии импульсов VNS составляла 0,5 секунды (рис. 5а). Каждая последовательность импульсов доставлялась примерно каждые 30 секунд в течение 2,5 часов.Ни в одном случае не было настроек, выходящих за рамки тех, которые используются для VNS при эпилепсии или депрессии (выходные токи были ≤3,5 мА, частоты ≤30 Гц, длительность импульса ≤1000 мкс и рабочие циклы (время включения / выключения) ≤50%.
Рисунок 5
( a ) Настройки стимуляции для парной терапии VNS. На нижней панели показаны настройки стимуляции (0,8 мА, 30 Гц), которые перекрываются с тоном. Каждая пара тонов VNS воспроизводилась каждые 30 секунд в течение примерно 2,5 часов за сеанс в течение 24 часов.( b ) Настройки стимуляции для непарной (контрольной) группы. Во время каждого сеанса участники получали только 10 минут тональных сигналов, 5 минут молчания и без VNS; 2 часа только VNS; 5 минут тишины и без VNS, а только 10 минут тонов.
В парной группе VNS каждый импульс VNS 0,5 с подавался одновременно с тональным сигналом 0,5 с каждые 30 с в течение 2,5 часов. Терапевтические тоны исключали одну или несколько частот шума в ушах участника. Тоны в паре с VNS находились на расстоянии не менее ½ октавы от наиболее заметной частоты шума в ушах для каждого отдельного участника.Частоты колебались от 170 до 16000 Гц. Звуки воспроизводились с интенсивностью, зависящей от уровня комфорта участника, с поправкой на любую потерю слуха на разных частотах, а интенсивность была ограничена до 80 дБ SPL. Каждая из частот тона должна казаться возникающей из различных трехмерных местоположений (запрограммированных с использованием модели головы KEMAR), чтобы избежать смещения при представлении тона (в паре с VNS) из одной пространственной точки. Частота и интенсивность (дБ SPL) каждого тона выбирались случайным образом каждый раз, когда подавался импульс VNS.
В контрольной (непарной) группе VNS не было сопряжено с тонами (только 10 минут тонов, 5 минут тишины и отсутствие VNS; 2 часа только VNS; 5 минут молчания и без VNS и 10 минут тонов) только) в течение 2,5 часов (рис. 5б).
Проведение терапии на дому
Обе группы получали терапию в течение 6 недель (рандомизированная часть исследования). Затем контрольная группа перешла к настройкам парной группы VNS через 6 недель, в то время как группа VNS продолжила исходную парную терапию VNS.
После того, как аудиолог запрограммировал настройки системы, участники обучили правильному использованию устройства для инициирования ежедневной стимуляции дома. Хотя стимуляция проводилась самим участником у себя дома, сеанс тестовой терапии проводился на месте под наблюдением исследователя. VNS вводили, чтобы определить, может ли участник перенести стандартные настройки и надлежащим образом начать терапию. Если участник не переносил стандартные настройки, выходной ток уменьшался с 0.8 мА с шагом 0,1 мА до достижения допустимого уровня. После получения приемлемого уровня выходного тока оператор объекта проверил, что участник может слышать тоны и что тона исходят из разных мест, как они воспринимаются через наушники (персонал объекта будет слушать тоны через наушники, а затем слушайте и убедитесь, что звуковые сигналы были слышны).
С участниками связались, по крайней мере, один раз в течение первых семи дней терапии, чтобы центр мог проверить, что участник проводил терапию надлежащим образом в течение всех 2 дней.5-часовая сессия. Согласие участников с проведением терапии на дому проверялось на местах путем просмотра файлов журнала данных (файлы журнала содержали информацию о дате / времени / стимуляции), чтобы можно было проверить ежедневное использование каждого участника.
Участников проинструктировали проводить терапию в тихой комнате и читать журнал или книгу, сидя в удобном кресле. Им также велели не спать, не смотреть телевизор (со звуком) и не вести длительные разговоры во время терапии. Однако им было разрешено работать на компьютере с отключенным звуком.Намерение состояло в том, чтобы позволить участнику слышать звуковые сигналы, но при этом иметь возможность выполнять некоторые другие задачи. Хотя участникам было дано указание проводить терапию у себя дома примерно в одно и то же время каждый день, им было разрешено проводить терапию в разное время из-за конфликтов в расписании. Если участникам требовался перерыв в туалете или возникали другие социальные проблемы (например, телефонный звонок), им давали 15-минутный перерыв. Кроме того, если участник пропускал в среднем более трех сеансов терапии в неделю или более трех сеансов подряд, этот участник исключали из первичного анализа, но включали в любой анализ намерения лечить.Проведение терапии проверялось путем просмотра записей на компьютере участника при каждом визите в исследование.
Базовый скрининг и показатели результатов
Базовый скрининг включал аудиометрию (до 12 кГц с чистыми тонами как для стимулов воздушной, так и для костной проводимости, включая тестирование на распознавание односложных слов) и сопоставление высоты тиннитуса. Пороги слуха были измерены при 0,5 кГц, 1 кГц, 2 кГц, 3 кГц, 4 кГц, 6 кГц, 8 кГц и 12,5 кГц (рис. 6). Оценки включали инвентаризацию шума в ушах (THI), анкету для инвалидов в ушах (THQ), функциональный индекс шума в ушах (TFI), степень тяжести (участников просили оценить громкость своего шума в ушах по шкале от 0 до 100; 0 означает отсутствие шума в ушах и 100 указывает на самый громкий шум в ушах, который они могут себе представить), а также показатели психоакустического исхода, включая минимальный уровень маскировки (MML) и соответствие громкости 48,49 .Также были получены данные о депрессии Бека (BDI) 50 , опроснике состояния и тревожности (STAI) 51 и SF-12 52 . Поскольку это было пилотное исследование, не было назначено никаких первичных критериев оценки результатов. Перед началом терапии у участников было три базовых визита. Первый визит был проведен вскоре после получения согласия для подтверждения права на участие. Второй был проведен как минимум через две недели. Затем была запланирована имплантация устройства, и участники были рандомизированы в парную группу VNS или контрольную группу (описано ниже).Третью базовую оценку проводили после операции, но до начала терапии, чтобы определить, оказали ли операция или анестезия какое-либо существенное влияние на оценки. Всем участникам оценивали результаты в клинике.
Рис. 6
Аудиометрия в чистом тоне как для парной VNS, так и для контрольной группы. Аудиограммы были получены на исходном уровне до имплантации. Пороги слуха были измерены при 0,5 кГц, 1 кГц, 2 кГц, 3 кГц, 4 кГц, 6 кГц, 8 кГц и 12.5 кГц.
Статистический анализ
Базовые характеристики отображались для каждой группы в виде среднего значения и стандартного отклонения для непрерывных переменных и чисел на категорию для категориальных переменных. Базовые характеристики сравнивались между группами с использованием t-критерия или критерия хи-квадрат. Отклонение от исходного уровня рассчитывалось для каждой группы как процентное или абсолютное изменение от исходного уровня до 6 недель. Изменение исхода внутри группы (THI) от исходного уровня до 6-й недели было проанализировано с помощью одновыборочного t-критерия (H 0 : изменение = 0) и представлено в виде средней разницы (с доверительным интервалом 95%) для каждого группа.Сравнения между группами проводились с помощью двухвыборочного t-критерия. Для всех сравнений альфа была установлена на 0,05. Никаких поправок на множественность не делалось, и каждый анализ был протестирован на уровне 0,05.
ANOVA с повторными измерениями выполняли с условием (парное VNS по сравнению с контролем) в качестве переменной между субъектами и THI (на исходном уровне, 6 недель и 12 недель) в качестве переменной внутри субъекта. Аналогичный анализ был проведен для THQ, TFI, уровня громкости, MML и соответствия громкости. Чтобы проверить, есть ли разница между двумя группами (парные vs.контроль) был равен, мы применили тест сферичности Мочли. Этот тест не показал статистически значимых различий между дисперсиями (Mauchly W = 0,84, χ 2 = 4,79, p = 0,091).
Также был проведен анализ респондентов для определения клинически значимых эффектов. Для THI снижение на 20% или абсолютное снижение на 7 пунктов считалось клинически значимым 29,30,52 . Снижение THQ на 10 пунктов и снижение TFI на 13 или 7 пунктов считалось клинически значимым 31,53 .Для сравнения респондеров и не отвечающих как для парной группы VNS, так и для контрольной группы, был рассчитан коэффициент χ 2 . Статистический анализ был выполнен с использованием MATLAB (Mathworks, Inc.), SPSS и SAS v 9.4.
Как создать быстрый и высокочувствительный звуковой извещатель, который остается устойчивым к температурным перепадам
Введение
Лягушки-быки, прыгающие в холодный пруд, должны быстро адаптироваться к резкому перепаду температуры. Действительно, лягушки приспосабливаются к широкому диапазону физиологических температур тела (Lillywhite, 1970).Летом самцы лягушек собираются вместе и хором привлекают самок (Capranica, 1965). Чтобы успешно найти партнеров для спаривания, их слух должен быть острым в теплые летние месяцы. Вокализация лягушки-быка состоит из звуков низкой и средней частоты (200–2000 Гц) (Capranica, 1965; Smotherman, Narins, 2000; Simmons, 2004). Кроме того, слух также важен для охоты на насекомых и территориального поведения (Emlen, 1968; Wiewandt, 1969). Насекомые машут крыльями с частотой от 330 Гц (комнатные мухи) до 600 Гц (комары) и даже могут достигать 1040 Гц (маленькие кусающие мошки).Все эти звуковые частоты улавливаются и передаются одним органом слуха лягушки — сосочком земноводных (van Dijk et al., 2011). Соответственно, записи спайков в волокнах слухового нерва сосочка земноводных лягушки отображают кривые настройки с наибольшей чувствительностью и лучшими частотами (или самыми низкими порогами уровня звука) ∼500–650 Гц (Feng et al., 1975; Heffner and Heffner, 2007).
Волосковые клетки преобразуют звуковые колебания в ступенчатые электрические сигналы, которые затем отправляются в мозг через пики потенциала действия (AP) в афферентных волокнах.При более высоких температурах, in vivo записи одного афферентного волокна показали увеличение частоты спонтанных спайков, снижение порога интенсивности звука, уменьшение задержки ответа на звук и более высокую векторную силу (или лучшую точность фазовой синхронизации) (Stiebler и Наринс, 1990; ван Дейк и др., 1990). Это указывает на то, что орган слуха лягушек передает больше звуковой информации с более высокой чувствительностью, более коротким временем реакции и большей временной точностью при более высоких температурах.Каковы клеточные и синаптические механизмы, которые объясняют эти наблюдения in vivo ?
Волосковые клетки обнаруживают и преобразовывают три аспекта звука: интенсивность, фазу и частоту. Информация о быстром возникновении и смещении звуковых переходных процессов также должна достоверно передаваться в слуховые нервы в синапсах ленточного типа (Rutherford, 2015; Coate et al., 2019). Действительно, волосковые клетки экспрессируют ионные каналы с одной из самых быстрых кинетик активации и деактивации (Engel, 2008; Heil and Peterson, 2017; Pangrsic et al., 2018). Звуковые сигналы передаются через токи трансдукции (I), опосредованные притоком K + в пучки стереоцилий, что приводит к постепенным изменениям мембранного потенциала рецептора (V m ). Обнаружение низкоуровневых звуков облегчается, если волосковые клетки имеют большое входное сопротивление (R в ), учитывая, что V m = R в × I. точность требует более коротких постоянных времени мембраны (τ м = R в × C м , где C м — емкость мембраны волосковой клетки), что требует небольшого R в .Как волосковая клетка справляется с этими противоречивыми требованиями к ее биофизическим свойствам? Уменьшается ли количество R в волосковой клетки при повышении температуры, как это наблюдается в других нейронах лягушки-быка (Santin et al., 2013)? Если да, то как слуховые волосковые клетки и их синапсы компенсируют температурно-зависимые изменения R в , чтобы поддерживать как звуковую чувствительность, так и точность воспроизведения во времени?
Чтобы ответить на эти вопросы, мы выполнили in vitro, записи напряжения и тока фиксации отдельных волосковых клеток и их афферентных волокон в сосочках земноводных лягушки-быка как в помещении (23–25 ° C), так и при высоком (30 ° C). C – 33 ° C) температура.Наши результаты показывают, что большие амплитуды и более быстрая кинетика тока Ca 2+ и K + приводят к более высоким собственным частотам электрического резонанса волосковых клеток, тогда как более короткие синаптические задержки, более синхронное мультивезикулярное высвобождение и снижение R в при высокой температуре вносят свой вклад. для более точной синхронизации по фазе звуковых сигналов. Более того, мы предполагаем, что волосковые клетки компенсируют более низкий уровень R в при высокой температуре за счет увеличения размера легко высвобождаемого пула (RRP) везикул и эффективности экзоцитоза, что приводит к повышению чувствительности к звуку.
Результаты
Большинство афферентных волокон генерируют больше спайков при более высокой температуре
Чтобы определить температурную зависимость спонтанных спайков на афферентных волокнах волосковых клеток, мы зарегистрировали спонтанные изменения потенциала в афферентных волокнах, используя токовые клещи на всю клетку с подачей нулевого тока. При 23,7 ± 0,2 ° C частота спайков составляла 2,0 ± 0,64 Гц, амплитуда составляла 51,9 ± 6,8 мВ, порог составлял -51,0 ± 2,0 мВ, а мембранный потенциал покоя (V rest ) афферентных волокон составлял -66.9 ± 2,1 мВ ( n = 18). Среди этих волокон семь из них показали большие выбросы, которые превышали 0 мВ с более высоким порогом (рис. 1 A1 ). Амплитуда этих больших всплесков составляла 85,7 ± 5,7 мВ ( n = 6), тогда как амплитуда выбросов без превышения составляла 35,0 ± 4,8 мВ ( n = 12, p <0,0001). Порог больших всплесков составлял -44,1 ± 2,4 мВ ( n = 6), тогда как порог малых всплесков составлял -54,8 ± 2,1 мВ ( n = 12, p = 0.0063, непарный т проба). Статистической разницы между этими двумя группами по частоте не было (превышение: 2,02 ± 0,56 Гц по сравнению с непревращением: 1,99 ± 0,94 Гц, p = 0,98, непарный тест t ) и V rest (превышение: -64,1 ± 4,6 мВ по сравнению с непревращением: -68,3 ± 2,3 мВ, p = 0,37).
Рис. 1.
Неоднородность температурной зависимости выступов афферентных волокон. A1 , Запись токовых клещей целой клетки афферентного волокна с вводом нулевого тока при 23 ° C.Можно четко различить всплески AP и EPSP. Пунктирная линия показывает 0 мВ. A2 , Пик, показанный в прямоугольнике с пунктирной линией в A1 , был увеличен во временной шкале. Мембранный потенциал покоя (V rest ) составлял -74 мВ, а порог AP составлял -58 мВ. A3 , Спонтанные спайки от того же афферентного волокна, показанные в A1 , были зарегистрированы при 32 ° C. A4 , Шипы, показанные в рамке с пунктирной линией в A3 , увеличены во временной шкале.V , остаток составлял -78 мВ. Пороги первого и второго AP составляли -66 и -63 мВ соответственно. Пять из семи афферентных волокон (71%) вызвали больше спайков при высокой температуре. B1 , Спонтанные спайки были зарегистрированы от другого афферентного волокна при 25 ° C. Пунктирная линия показывает 0 мВ. B2 , Пик, показанный в прямоугольнике с пунктирной линией в B1 , был увеличен во временной шкале. V rest составлял -58 мВ, а порог AP составлял -46 мВ. B3 , Спонтанные выбросы из того же волокна, показанного в B1 , были зарегистрированы при 32 ° C. B4 , Пик, показанный в прямоугольнике с пунктирной линией в B3 , был увеличен во временной шкале. V rest составлял -60 мВ, а порог AP составлял -46 мВ. Два из семи афферентных волокон (29%) вырабатывали меньше шипов при высокой температуре.
Для 7 из этих 18 волокон мы успешно зарегистрировали спайки как при комнатной, так и при высокой температуре.Пять из них выстрелили больше шипов при высокой температуре (Таблица 1; Рис. 1 A1 – A4 ). Мы назвали эти волокна «термоположительными волокнами». Температурный коэффициент (Q 10 ) скорости спонтанного всплеска составлял 4,0 ± 1,2 ( n = 5), что было аналогично тому, что обнаружено в волокнах крыс (медиана Q 10 = 6,6) (Wu et al., 2016 ). Хотя порог срабатывания AP при высокой температуре (-58,6 мВ) был ниже, чем при комнатной температуре (-53,9 мВ, n = 5, p = 0.0158), уровни вероятности всплесков остаются неизменными при изменении температуры: при комнатной температуре 25,8% ВПСП вызвали всплеск; а при высокой температуре 25,4% ВПСП вызвали всплеск ( n = 5, p = 0,97). Уменьшение входного сопротивления мембраны афферентного волокна при высокой температуре приводит к более быстрому распаду ВПСП (см. Рис. 4), что затрудняет их суммирование, что противодействует потенциальному увеличению вероятности выброса, вызванному более низкими пороговыми значениями. Напротив, в нейронах спиральных ганглиев улитки млекопитающих 80–97% ВПСП вызывают спайк при комнатной температуре (Rutherford et al., 2012). Низкая частота отказов шипов EPSPs в волокнах млекопитающих может быть результатом меньшей площади поверхности и высокого входного сопротивления постсинаптического бутона млекопитающих (Rutherford et al., 2012). В сосочке земноводных лягушки-быка каждая волосковая клетка соединяется с 3–6 большими когтеобразными концами афферентных волокон, и каждое волокно получает несколько ленточных синапсов (Graydon et al., 2014), в то время как каждое афферентное волокно млекопитающих соединяется только с одной единственной синаптической лентой ( Либерман, 1980; Резерфорд, 2015).
Таблица 1.
Температурная зависимость шипов афферентных волокон
Два других волокна генерировали меньше шипов при высокой температуре (таблица 1; рис. 1 B1 – B4 ), и мы назвали эти «термонегативные волокна». Термонегативные волокна имели низкую частоту спонтанных импульсов (<1 Гц; Таблица 1). В слуховом нерве крыс частота спонтанных импульсов увеличивается во время созревания улитки с 3,87 Гц на P15 – P17 до 12,85 Гц на P29 – P32 (Wu et al., 2016). Возможно, что некоторые волокна с низкочастотными шипами у лягушки могут получать синаптическое высвобождение из некоторых незрелых волосковых клеток на краю сосочка амфибий (Lewis and Li, 1975).
Частоты спонтанных EPSC и EPSP усиливаются при высокой температуре
Чтобы изучить, как температура влияет на спонтанное высвобождение волосковых клеток, мы зарегистрировали спонтанные EPSC из афферентных волокон без ограничения напряжения их пресинаптических волосковых клеток под комнатой (черные следы) и высокой (красные следы) следы) температуры (рис.2 A1 , B1 ). Частота ЭПСК увеличилась с 78,2 ± 9,5 Гц при комнатной температуре до 115,2 ± 9,7 Гц при высокой температуре ( n = 18, p = 0.0097; Таблица 2). Распределение амплитуды EPSC афферентного волокна на рис. 2 A2 также показывает, что больше EPSC происходит при высокой температуре. Распределение амплитуды EPSC хорошо соответствовало функции Гаусса (рис.2 A2 : R 2 = 0,96 при 22 ° C; R 2 = 0,93 при 32 ° C). Пики гауссовой аппроксимации приходятся на 93,5 пА при 22 ° C и 89,7 пА при 32 ° C (рис. 2 A2 ), что позволяет предположить, что средняя амплитуда EPSC не увеличивается при высокой температуре.Большинство афферентных волокон (12 из 18) не показывали увеличения амплитуды EPSC при высокой температуре, а среднее значение амплитуды EPSC оставалось неизменным при изменении температуры ( p = 0,61; Таблица 2). На рис. 2 A3 средние значения EPSC были нормированы на их пиковые значения, и одна экспоненциальная функция использовалась для подбора фазы распада EPSC и определения постоянной времени затухания (τ распад ). Время нарастания 10–90% и затухание τ EPSC составило 223 ± 13 мкс и 617 ± 36 мкс ( n = 18; таблица 2) при комнатной температуре, что быстрее, чем измеренное в афферентных волокнах незрелых крыс. (Йи и др., 2010), но аналогично измеренным в афферентных волокнах взрослых черепах (Schnee et al., 2013) и слышащих крыс (Grant et al., 2010). Повышение температуры уменьшило время нарастания на 10–90% и распад τ EPSC со значениями Q 10 , равными 1,7 и 1,8, соответственно (Таблица 2). Это согласуется с нашими предыдущими находками, показывающими, что снижение температуры с 25 ° C до 15 ° C замедляет активацию и распад EPSC в афферентных волокнах лягушки-быка (Li et al., 2009). Точно так же в амакриновых клетках AII крысы повышение температуры с 26 ° C до 34 ° C уменьшает как время нарастания, так и τ распад mEPSCs со значениями Q 10 , равными 1.3 и 1.5 соответственно (Veruki et al., 2003). Вычисляя интегралы EPSC, мы обнаружили, что постсинаптический перенос заряда снижается при высокой температуре (таблица 2), что может быть связано с более быстрой активацией и дезактивацией (рис.2 A3 , B3 ) постсинаптических рецепторов AMPA и более быстрыми темпами. активность переносчика глутамата при высокой температуре (Auger and Attwell, 2000).
Рисунок 2.
Самопроизвольное увеличение частоты EPSC при высокой температуре. A1 , EPSC были записаны с афферентного волокна, которое было зафиксировано напряжением -90 мВ при 22 ° C (черный) и 32 ° C (красный). A2 , Всего было получено 6321 и 14451 EPSC при 22 ° C (черный) и 32 ° C (красный) соответственно. Распределения амплитуды EPSC были подогнаны с помощью функции Гаусса. A3 , Средние значения EPSC, записанные при 22 ° C (черный) и 32 ° C (красный), были нормализованы к их пиковому значению. Одноэкспоненциальная аппроксимация фазы спада усредненных EPSC показала, что τ decay составлял 0,46 мс при 22 ° C (черная пунктирная линия) и 0,40 мс при 32 ° C (красная пунктирная линия), соответственно. B1 , EPSC были записаны от другого афферентного волокна при 23 ° C (черный) и 31 ° C (красный). B2 , Всего было получено 2931 и 5041 EPSC при 23 ° C (черный) и 31 ° C (красный), соответственно. На распределении амплитуды присутствуют два пика. EPSC были разделены на две группы в зависимости от их размера: маленькие EPSC с амплитудой <150 пА и большие EPSC с амплитудой> 150 пА. Распределение амплитуды EPSC в каждой группе соответствовало функции Гаусса. B3 , Средние значения EPSC, записанные при 23 ° C (черный) и 31 ° C (красный), были нормализованы к их пиковому значению. Одноэкспоненциальная аппроксимация фазы спада усредненных EPSC показала, что τ decay составлял 0,47 мс при 24 ° C (черная пунктирная линия) и 0,26 мс при 31 ° C (красная пунктирная линия), соответственно.
Таблица 2.
Биофизические свойства EPSC и EPSP афферентных волокон
Среди всех зарегистрированных афферентных волокон 5 из 18 (28%) показали двойные пики в распределении амплитуды EPSC как при комнатной, так и при высоких температурах.На рисунке 2 B1 показаны EPSC, зарегистрированные от афферентного волокна, которое имело распределение с двумя пиками (рис. 2 B2 ). Двух- или трехпиковые распределения амплитуды EPSC наблюдались ранее в афферентных волокнах зрелых крыс (Grant et al., 2010) и черепах (Schnee et al., 2013). Мы разделили EPSC на две группы в зависимости от их амплитуды: маленькие EPSC с амплитудой <150 пА и большие EPSC с амплитудой> 150 пА. Частотные распределения амплитуды EPSC в каждой группе хорошо соответствовали функции Гаусса (небольшие события: R 2 = 0.94 при 23 ° C, R 2 = 0,93 при 31 ° C; большие события: R 2 = 0,93 при 23 ° C, R 2 = 0,97 при 31 ° C). При температуре ниже 23 ° C первый и второй пик гауссовой аппроксимации составляли 78 и 292 пА соответственно. При температуре ниже 31 ° C первый и второй пик составили 89 и 279 пА соответственно. При температуре ниже 23 ° C было 65% EPSC> 150 pA. Группа крупных EPSC увеличилась до 74% при 31 ° C.
Спонтанные ВПСП также регистрировались от афферентных волокон с использованием токовых клещей для всей клетки с током удержания 0 пА.В соответствии с EPSC частота EPSP увеличилась с 40,4 Гц при 23,4 ° C до 56,9 Гц при 31,5 ° C ( n = 19, p = 0,003; Таблица 2). Однако реакция амплитуд ВПСП была различной: 58% (11 из 19) волокон показали уменьшенную среднюю амплитуду ВПСП при высокой температуре. Это может быть вызвано сильной активацией токов K + в афферентах, которые притупляют амплитуду ВПСП при более высоких температурах. Однако усреднение всего набора данных не выявило статистической разницы в амплитуде ВПСП между комнатной и высокой температурой (3.9 ± 0,3 мВ против 3,8 ± 0,4 мВ соответственно; n = 19, p = 0,8; Таблица 2).
Рисунок 3 A показывает один пример с уменьшенной амплитудой EPSP при высокой температуре, а Рисунок 3 B показывает другой пример с увеличенной амплитудой EPSP при высоких температурах. Распределение амплитуды ВПСП показано на Рисунке 3 A2 , где было подсчитано 1250 и 1817 событий ВПСП от афферентного волокна при 23 ° C и 31 ° C соответственно. Амплитудные распределения ВПСП хорошо соответствовали функции Гаусса (рис.3 A2 : R 2 = 0,89 при 23 ° C; R 2 = 0,97 при 31 ° C). Пики гауссовой аппроксимации составляют 5,7 мВ при 23 ° C и 2,7 мВ при 31 ° C. Одноэкспоненциальная аппроксимация фазы распада усредненных ВПСП (рис. 3 A3 ) показала, что распад τ составлял 2,7 ± 0,4 мс при комнатной температуре ( n = 19; Таблица 2). Это быстрее, чем в афферентных волокнах незрелых крыс (Yi et al., 2010). EPSP, показанные на рисунке 3 B1 , были зарегистрированы с того же афферентного волокна с двухпиковым распределением амплитуды EPSC, как показано на рисунке 2 B2 .В течение 30-секундных записей было зарегистрировано 1054 и 1581 ВПСП от другого афферентного волокна при комнатной и высокой температуре соответственно (рис. 3 B2 ). Частотное распределение амплитуд ВПСП, показанное на рисунке 3 B2 , также имело двойные пики при высокой температуре, но не при комнатной температуре. При температуре 23 ° C пик гауссовой аппроксимации составил 2,7 мВ ( R 2 = 0,87). При температуре ниже 31 ° C первый и второй пик составили 3,4 мВ ( R 2 = 0.85) и 7,3 мВ ( R 2 = 0,81) соответственно. Кроме того, остаток афферентного волокна V был слегка деполяризован от -74,0 ± 0,7 мВ при 23 ° C до -71,2 ± 0,8 мВ при 32 ° C ( p = 0,0012, n = 19).
Рисунок 3.
Спонтанные ВПСП быстрее, и их частота увеличивается при высокой температуре. A1 , ВПСП были записаны с афферентного волокна при 23 ° C (черный) и 31 ° C (красный). Ток через мембрану волокна был ограничен 0 пА. A2 , Всего было зарегистрировано 1250 и 1817 EPSP при 23 ° C (черный) и 31 ° C (красный), соответственно. Распределения амплитуды ВПСП были аппроксимированы функцией Гаусса. A3 , Средние значения EPSP, зарегистрированные при 23 ° C (черная сплошная линия) и 31 ° C (красная сплошная линия), были нормализованы к их пиковым значениям. Пунктирные линии соответствуют одноэкспоненциальной фазе спада средних ВПСП. В этом примере τ распад составил 1,8 мс при 23 ° C (черная пунктирная линия) и 1.1 мс при 31 ° C (красная пунктирная линия) соответственно. B1 , ВПСП были записаны с того же афферентного волокна, как показано на рисунке 2 B1 – B3 , при 23 ° C (черный) и 31 ° C (красный). B2 , Всего было зарегистрировано 1054 и 1581 ВПСП при комнатной и высокой температуре. B3 , Средние значения EPSP, записанные при 23 ° C (черный) и 31 ° C (красный), были нормализованы к их пиковым значениям. Одноэкспоненциальная аппроксимация фазы спада усредненных ВПСП (пунктирные линии) показала, что τ распад равен 1.1 мс при 23 ° C и 0,7 мс при 31 ° C соответственно.
Входное сопротивление мембраны афферентного волокна при высокой температуре
Распад τ ВПСП уменьшился при высокой температуре во всех зарегистрированных волокнах (Рис. 3 B3 ; Таблица 2), что может быть связано с более низким входным сопротивлением мембраны ( R в ) при высокой температуре. Таким образом, мы оценили R в афферентных волокнах с использованием токовых клещей: R в = ΔV / ΔI, где ΔV было изменением мембранного потенциала, вызванным инжекцией тока 100 пА (рис.4 А ). R в афферентных волокнах снизился с 557 ± 128 мОм при 22,0 ± 0,1 ° C до 395 ± 85 мОм при 32,0 ± 0,2 ° C ( n = 5, p = 0,027). В этой группе волокон τ распад ВПСП составил 6,5 ± 2,7 мс и 3,3 ± 1,7 мс при комнатной и высокой температуре соответственно ( n = 5, p = 0,041). Распад τ положительно коррелировал с афферентным волокном R в (наклон линейной регрессии = 0,018 ± 0,003, R 2 = 0.79, p. = 0,0006; Рис.4 B , синяя линия).
Рис. 4.
Входное сопротивление мембраны афферентного волокна и кинетика затухания ВПСП. A , В токовых клещах афферентные волокна были гиперполяризованы путем введения отрицательных токов от -50 до -150 пА в течение 100 мс. Средние значения реакции мембранного потенциала одного афферентного волокна при 22 ° C и 31 ° C показаны черным и красным цветом соответственно. Красные и черные стрелки указывают на абсолютное изменение мембранных потенциалов при 22 ° C и 31 ° C соответственно.Входное сопротивление афферентного волокна рассчитывалось по закону Ома, R в = ΔV / ΔI. Фаза спада мембранного потенциала описывалась двойной экспоненциальной функцией (серые пунктирные линии). B , τ распад ВПСП был нанесен на график против R в соответствующих волокон. Каждая пара точек данных, записанных с одного и того же афферентного волокна, соединена пунктирной линией: комнатная (черная) и высокая (красная) температура ( n = 5). Подгонка линейной регрессии показана синим цветом и указывает на положительную корреляцию между EPSP τ , распад и R в (наклон = 0.018 ± 0,003, R 2 = 0,79, p = 0,0006).
Афферентное волокно состоит из двух отсеков: окончания типа чашечки, контактирующего с волосковой клеткой, и длинного тонкого волоконного кабеля. Таким образом, мы аппроксимируем фазу спада изменения мембранного потенциала ступенчатым током с двойной экспоненциальной функцией (серые пунктирные линии наложены на кривую V м ; рис. 4 A , серые пунктирные линии наложены на кривую V м ) как следует:
Затем мы можем рассчитать средневзвешенную постоянную времени τ среднее значение , используя следующее:
Значение τ , среднее значение было короче при высокой температуре (4.8 ± 1,5 мс), чем при комнатной температуре (7,8 ± 1,5 мс, n = 5, p = 0,0014). Τ 1 при комнатной и высокой температуре составляло 3,2 ± 1,2 мс и 2,5 ± 1,5 мс соответственно, а τ 2 при комнатной и высокой температуре составляло 10,3 ± 1,3 мс и 6,0 ± 1,7 мс соответственно. Тот факт, что для соответствия откликам напряжения требуется двойная экспоненциальная функция с быстрой (τ f ) и медленной (τ s ) постоянными времени, указывает на то, что афферентное волокно не является изопотенциальным (Golowasch et al., 2009), в отличие от волосковой клетки (см. Рис. 6). Поскольку афферентный аксон имеет обрезанный конец из-за процесса изоляции сосочка амфибии, его можно смоделировать как конечный кабель с электротонической длиной, определяемой следующим образом (Rall, 1969):
Таким образом, мы находим, что L = 2,1 для 22 ° C и L = 2,6 для 32 ° C для афферентных волокон. Это относительно большое значение по сравнению с нейронами гиппокампа, где L = 0,9 (Brown et al., 1981), что свидетельствует о сильном ослаблении напряжения от конца постсинаптической чашечки до кончика отсеченного и запечатанного афферентного аксона.Следовательно, кластер каналов Na + на первой аксональной геминоде д. Быть довольно близко к концу чашечки, чтобы запустить спайк (Kim and Rutherford, 2016). Напротив, амплитуда ВПСП не изменилась значительно с температурой для этого набора данных (6,5 ± 0,7 мВ против 9,3 ± 2,2 мВ при 22,1 ± 0,2 ° C и 32,0 ± 0,1 ° C, соответственно; n = 5, p = 0,3 ). Линейная регрессия амплитуды ВПСП и R в также не показала значимой разницы (наклон = -0.0016 ± 0,005, R 2 = 0,013, p = 0,76). В заключение, R в афферентных волокон демонстрирует значительную неоднородность от 100 мОм до 1000 мОм, что может способствовать изменению in vivo частоты спонтанных спайков и порогов афферентных волокон к звуковой стимуляции (Stiebler and Narins, 1990; Shrestha и др., 2018).
Изменения R в в зависимости от температуры также важны из-за соображений шума мембранного потенциала (Fatt and Katz, 1952; Johnson, 1987).Среднее квадратичное напряжение мембраны определяется теоремой Найквиста следующим образом:
здесь k B — постоянная Больцмана, T — температура в градусах Кельвина (T = 273 + температура в градусах Цельсия), R в — входное сопротивление мембраны, R a — осевое сопротивление кабеля и Δ f — полоса частот шума. Ширина полосы частот обратно пропорциональна постоянной времени мембраны, которая определяет частоту отсечки ячейки, и, таким образом, будет увеличиваться при более высоких температурах.Уменьшение R в с повышением температуры, таким образом, важно, потому что оно противодействует увеличению ширины полосы шума с температурой. Это помогает поддерживать мембранный потенциал на стабильном подпороговом уровне.
Мембранный потенциал покоя волосковых клеток
Увеличение частоты EPSC при высокой температуре предполагает более спонтанное высвобождение пузырьков из волосковых клеток, что может быть связано с деполяризацией V покоя при высокой температуре. Таким образом, мы измерили волосковую ячейку V rest под токовыми клещами для всей ячейки с током удержания на уровне 0 пА.Неожиданно записи целых клеток показали, что остаток V гиперполяризован от -65,0 ± 1,8 мВ при 23,2 ± 0,3 ° C до -70,1 ± 2,3 мВ при 31,2 ± 0,3 ° C ( p = 0,006, n = 15; Рис.5 А , черные кружки). Чтобы предотвратить диализ составляющих волосковых клеток через пипетку с пластырем, мы также выполнили записи с перфорированными пластырями с использованием грамицидина (40–50 мкг / мл). Перфорированный патч-зажим показал, что V rest не изменился при высоких температурах (V rest = -62.8 ± 1,3 мВ при 23,4 ± 0,5 ° C и V покой = -62,7 ± 1,5 мВ при 31,6 ± 0,2 ° C, p = 0,88, n = 9; Рис.5 A , синие точки).
Рис. 5.
Мембранный потенциал покоя волосковой клетки и ионные токи при высокой температуре. Раствор внутренней пипетки на основе A , A K + и записи токовых клещей с вводом нулевого тока использовались для измерения V остатка волосковых клеток. Записи всех клеток показали, что остаток V гиперполяризован при высокой температуре (черные кружки, p = 0.006, № = 15). Записи перфорированных пластырей, опосредованных грамицидином, показали, что V rest оставался таким же при высокой температуре (синие точки, p = 0,88, n = 9). B , Записи с фиксацией напряжения целых клеток волосковых клеток с внутренним раствором на основе K + показали изменения тока в ответ на ступенчатую деполяризацию от -90 мВ до -60 мВ. Следы усредняли для пяти волосковых клеток, записанных при 22 ° C (черный) и 32 ° C (красный). Обратите внимание на более быструю кинетику токов при 32 ° C. C , Чтобы выявить ток Ca 2+ , волосковые клетки закрывали внутренним раствором на основе Cs + -TEA. Волосковые клетки деполяризовались с линейной деполяризацией от -90 мВ до -50 мВ в течение 400 мс при 22 ° C (черный) и 31 ° C (красный). Следы усредняли по пяти волосковым клеткам. D , Зависящие от напряжения токи K + и Ca 2+ запускались одним и тем же стимулом линейного изменения с внутренним раствором на основе K + . Следы усредняли для 14 волосковых клеток, записанных при 22 ° C (черный) и 32 ° C (красный).Обратите внимание, как большие направленные наружу токи K + доминируют на кривой I – V волосковых клеток. ** p <0,01.
Калийные и кальциевые токи волосковых клеток при высокой температуре
Зажим при пониженном напряжении и с помощью внутреннего пипеточного раствора на основе K + мы определили зависящие от напряжения токи, которые активируются ступенчатой деполяризацией от -90 до -60 мВ. За быстро активирующимся входящим током последовал быстрый и большой выходящий ток (рис. 5 B ).Входящий ток представлял собой ток Ca 2+ , который быстрее активируется при более высоких температурах в волосковых клетках лягушки (Hudspeth and Lewis, 1988). Используя внутренние растворы на основе Cs + -TEA или K + , мы записали кривые I – V для токов Ca 2+ (рис. 5 C ) и всего K + . + Ca 2+ токов (рис. 5 D ) соответственно. Оба типа токов увеличиваются по амплитуде при высокой температуре. Важно отметить, что наклон зависимого от напряжения тока Ca 2+ был больше при высокой температуре, что делало приток Ca 2+ более чувствительным к небольшим изменениям мембранного потенциала.Увеличение тока целых клеток наружу K + + Ca 2+ при высокой температуре объясняет уменьшение V rest с температурой (рис. 5 A , черные кружки). В перфорированных патч-записях отсутствие изменения в V rest может быть связано с одновременным и более компенсирующим увеличением входящего тока Ca 2+ и наружного тока K + при повышении температуры.
Входное сопротивление мембраны волосковой клетки уменьшается при высокой температуре
При высокой температуре увеличение токов Ca 2+ и K + вокруг V остального должно приводить к уменьшению входного сопротивления мембраны (R в ).Чтобы проверить это, мы выполнили регистрацию напряжения всей клетки и токовые зажимы волосковых клеток. В зажиме напряжения гиперполяризованные волосковые клетки от -90 мВ до -95 мВ или -100 мВ в течение 10 мс вызвали небольшие внутренние токи (рис. 6 A1 ). R в был рассчитан по закону Ома (R в = ΔV / ΔI). R в волосковых клетках уменьшился при высокой температуре (659,7 ± 54,3 мОм при 23,2 ± 0,2 ° C и 343,7 ± 34,7 мОм при 31,4 ± 0,2 ° C, p = 0,0006, n = 7; Рис.6 A2 ).В условиях токового зажима инъекция -20 пА или -30 пА гиперполяризовала клетки (фиг. 6 B1 ). Токовые клещи также показали уменьшение R в при высокой температуре (640,3 ± 56,4 мОм при 23,2 ± 0,2 ° C и 389,3 ± 28,2 мОм при 31,2 ± 0,2 ° C, p = 0,0023, n = 7; рис. 6 B2 ). Изменения тока или мембранного потенциала определяли путем расчета разницы между базовой линией и плато (рис. 6 A1 , пунктирные линии, B1 ). Не было статистически значимой разницы в R и , измеренных с помощью токовых клещей или токовых клещей в любой комнате ( p = 0.2598) или высокой ( p = 0,7016) температуры ( n = 7).
Рис. 6.
Влияние температуры на свойства пассивной мембраны волосковых клеток. A1 , Входящие токи были вызваны небольшой гиперполяризацией от -90 до -100 мВ при 23 ° C (черный) и 31 ° C (красный). Величина тока представляла собой разницу между базовой линией и внутренним током плато (пунктирные линии). B1 , Гиперполяризованный мембранный потенциал в ответ на подачу тока -30 пА под токовыми клещами регистрировали при 23 ° C (черный) и 31 ° C (красный).Изменение мембранного потенциала рассчитывали путем вычитания базовой линии из плато (пунктирные линии). A2 , Используя измерения напряжения и R в = ΔV / ΔI, входное сопротивление мембраны (R в ) было ниже при высокой температуре ( p = 0,0006, n = 7). B2 , Измерения токового зажима также показали, что R в уменьшился при высокой температуре ( p = 0,0031, n = 7). B3 , Постоянная времени мембраны (τ) была оценена путем подгонки одной экспоненты к временному окну 40 мс отклика напряжения после подачи начального тока ( B1 , наложенные синие пунктирные линии).τ было быстрее при высокой температуре ( p <0,0001, n = 7). B4 , C m , рассчитанный с использованием C m = τ / R в , не показал значительных изменений при высокой температуре ( p = 0,2506, n = 7). A2 , B2 – B4 , точки представляют отдельные значения; белые кружки обозначают среднее значение; столбцы представляют SEM. ** p <0,01, *** p <0,001, **** p <0.0001.
Некоторые волосковые клетки имеют тенденцию быть более протекающими к концу регистрации целых клеток, и это может повлиять на R в (Ceballos et al., 2017). Чтобы контролировать эту возможность, мы сначала записали R в при комнатной температуре, а затем нагрели раствор до высокой температуры. R в снизился с 578,8 ± 49,8 мОм при 24,0 ± 0,2 ° C до 315,8 ± 33,3 мОм при 31,5 ± 0,2 ° C ( p = 0,0001, n = 8). Затем мы записали данные из отдельной группы клеток, сначала при высокой температуре, а затем при комнатной температуре.R в также был ниже при высокой температуре (600,2 ± 38,2 мОм при 24,1 ± 0,1 ° C и 277,6 ± 34,0 мОм при 31,9 ± 0,1 ° C, p = 0,0146, n = 3). Таким образом, порядок записи не повлиял на R в , измеренных при комнатной ( p = 0,81) или высокой ( p = 0,53) температуре. Это говорит о том, что снижение R в при высокой температуре произошло не из-за того, что волосковые клетки стали более протекающими к концу записи, а из-за увеличения проводимости клеточной мембраны при высокой температуре (рис.5 В ).
Используя данные токовых клещей, мы также рассчитали постоянную времени мембраны (τ). Мы установили одну экспоненту для временного окна 40 мс отклика напряжения после первоначальной подачи тока (рис. 6 B1 , наложенная синяя пунктирная линия). Постоянная времени составляла 8,50 ± 0,73 мс при 23,2 ± 0,2 ° C и 5,67 ± 0,61 мс при 31,2 ± 0,2 ° C ( p <0,0001, n = 7; рис. 6 B3 ). Затем мы определили емкость мембраны (C м ), используя уравнение C м = τ / R в .Среднее значение C m оставалось неизменным независимо от изменений температуры (13,5 ± 0,9 пФ при 23,2 ± 0,2 ° C и 14,6 ± 1,2 пФ при 31,2 ± 0,2 ° C, p = 0,2506, n = 7; Рис.6 B4 ). Это значение C m похоже на предыдущие записи для целых клеток при ограничении напряжения C m в волосковых клетках лягушки-быка (Li et al., 2009). Такое согласие между значениями напряжения и тока фиксации для R в и C m ожидается только для изопотенциальных ячеек (Golowasch et al., 2009).
Электрическая резонансная частота увеличивается при высокой температуре
В отличие от механической настройки в улитке млекопитающих волосковые клетки сосочка амфибий у лягушек настраиваются электрически (Smotherman and Narins, 2000). Электрический резонанс волосковых клеток сосочка земноводных соответствует характеристическим (лучшим) частотам афферентных волокон (Smotherman, Narins, 1999). Электрический резонанс является результатом взаимодействия потенциалзависимых ионных каналов и мембранной емкости (Crawford and Fettiplace, 1981; Art and Fettiplace, 1987; Hudspeth and Lewis, 1988).Поскольку проводимость потенциалзависимых кальциевых и калиевых каналов зависит от температуры, как показано на рисунке 5, мы ожидаем, что электрический резонанс будет чувствительным к температуре. Чтобы вызвать электрическое кольцо вокруг потенциалов покоя мембраны, волосковым клеткам вводили ступенчатый ток от нуля под цельной клеткой (фиг. 7 A ) или перфорированным патч-зажимом (фиг. 7 B ). Резонансная частота была оценена путем подбора первых 20 мс затухающих колебаний мембранного напряжения с помощью синусоидальной волновой функции (Li et al., 2014). В предыдущем исследовании (Smotherman and Narins, 1999) было показано, что резонансная частота увеличивается по мере того, как к ячейке прикладываются большие шаги деполяризующего тока. Соответственно, деполяризующие волосковые клетки с шагом тока 100 пА вызывают резонансную частоту 236 ± 26 Гц, тогда как шаги тока 200 пА вызывают более высокую резонансную частоту 283 ± 31 Гц при комнатной температуре ( p = 0,0038, n = 4 ; Рис.7 A , C ). Чтобы сравнить резонансную частоту, измеренную с записями целых клеток, с записями с перфорированными заплатами, мы выбрали большие шаги тока 200 пА.Резонансная частота увеличилась с 283 ± 31 Гц при 21,8 ± 0,4 ° C до 472 ± 56 Гц при 31,4 ± 0,6 ° C ( p = 0,017, n = 4; рис.7 D , черный кружок). Аналогичным образом, записи перфорированных участков показали, что резонансная частота увеличилась с 362 ± 50 Гц при 24,8 ± 2,1 ° C до 597 ± 19 Гц при 31,5 ± 0,3 ° C ( p = 0,025, n = 3; Рис.7 D , синие точки). Не было статистической разницы в резонансной частоте, измеренной с помощью записи целых клеток и перфорированного пластыря в любой комнате ( p = 0.22, непарный t тест) или высокие ( p = 0,13, непарный t тест) температуры.
Рисунок 7.
Резонансная частота волосковой клетки увеличивается при высокой температуре. A , Электрический резонанс был вызван стимуляцией волосковых клеток с шагом тока в области целых клеток. Черные и красные следы были вызваны шагом тока 200 пА при комнатной и высокой температуре соответственно. Серый след вызван шагом тока 100 пА при комнатной температуре. Каждая из этих трех трасс представляет собой средний ответ трех волосковых клеток.Резонансные частоты были рассчитаны путем подгонки первых 20 мс изменений мембранного напряжения с помощью синусоидальной волновой функции (вставка из пунктирной линии). Резонансные частоты черных, красных и серых следов составляли 314, 517 и 262 Гц соответственно. B , Электрический резонанс волосковых клеток был вызван ступенью тока 200 пА в перфорированном пластыре, опосредованном грамицидином. Черные и красные следы были усреднены для 3 клеток, записанных при комнатной и высокой температуре соответственно. Резонансные частоты черных и красных следов составляли 356 и 611 Гц соответственно. C , Резонансные частоты волосковых клеток, вызванные шагами тока 100 пА, были ниже, чем частоты, вызванные шагами тока 200 пА ( n = 4, p = 0,004). D , Результаты как цельноклеточных (черный, n = 4, p = 0,017), так и перфорированных (синий, n = 3, p = 0,025) записей показали, что резонансная частота волосковых клеток значительно увеличивается при высокой температуре. Статистической разницы между резонансными частотами, измеренными с помощью цельноклеточного пластыря или перфорированного пластыря в любой комнате, не было ( p = 0.80, непарный t тест) или высокая ( p = 0,56, непарный t тест) температура. * p <0,05, ** p <0,01.
Резонансная частота волосковых клеток также увеличивается, когда мембранный потенциал покоя деполяризуется (Hudspeth and Lewis, 1988; Smotherman and Narins, 1999). Однако мембранный потенциал покоя не изменился для записей целых клеток на Фигуре 7 A (-65,2 ± 0,5 мВ при комнатной температуре против -63,8 ± 0,8 мВ при высокой температуре, p = 0.23, n = 4). Точно так же потенциал покоя мембраны не изменялся с температурой для данных перфорированного пластыря (-66,0 ± 2,7 мВ при комнатной температуре против -65,2 ± 2,6 мВ при высокой температуре, p = 0,51, n = 3). Следовательно, увеличение резонансной частоты при высокой температуре не связано с изменениями мембранного потенциала покоя. Вместо этого они вызваны более быстрой активацией и большей проводимостью каналов Ca 2+ и K + при деполяризации при высокой температуре (Hudspeth and Lewis, 1988; Smotherman and Narins, 1999).Резонансная частота, измеренная при комнатной температуре, соответствует результатам in vitro записи целых клеток волосковых клеток из сосочка земноводных с использованием протокола ZAP (Frolov and Li, 2017). Q 10 резонансной частоты волосковых клеток сосочка земноводных (1,7 ± 0,1, n = 4 для целой клетки; 1,9 ± 0,08, n = 3 для перфорированного участка) аналогичны значениям, зарегистрированным для мешковидных волосковых клеток лягушки. (Смотерман, Наринс, 1998). Этот зависящий от температуры сдвиг резонансной частоты волосковых клеток может быть ответственным за вызванные температурой сдвиги наилучшей частоты, наблюдаемые на in vivo записях слуховых нервных волокон сосочка амфибий (Stiebler and Narins, 1990).
Эффективность высвобождения увеличивается при высокой температуре
Более низкий уровень R в при высокой температуре приводит к меньшим изменениям мембранного потенциала, что снижает чувствительность волосковых клеток к звукам низкого уровня. Чтобы выяснить, компенсируют ли волосковые клетки эту предполагаемую потерю чувствительности, мы изучили температурную зависимость экзоцитоза пузырьков из волосковых клеток, используя измерения емкости мембран с временным разрешением (Lindau and Neher, 1988; Gillis, 2000). Изменения C m (ΔC m ) были вызваны ступенями деполяризации напряжения.Сначала мы использовали шаги деполяризации 20 мс от удерживающего потенциала от -90 до -30 мВ, чтобы вызвать быстрое высвобождение, которое истощает RRP, состоящий из пузырьков, пристыкованных к сгруппированным каналам Ca 2+ (Graydon et al., 2011) (рис. 8 A1 ). Деполяризация вызвала больший ток Ca 2+ (рис.8 A1 ) при высокой температуре, чем при комнатной температуре ( p <0,0001, n = 7): перенос заряда Ca 2+ (Q ) Ca ) было 11.9 ± 1,3 пКл при 23,3 ± 0,2 ° C и 15,9 ± 1,7 пКл при 32,0 ± 0,3 ° C. Q 10 вызванного импульсом 20 мс Q Ca составлял 1,5 ± 0,05 ( n = 7). Средняя емкость мембраны в состоянии покоя, измеренная при высокой температуре (16,9 ± 0,7 пФ), была аналогична измеренной при комнатной температуре (15,7 ± 0,6 пФ, p = 0,0841, n = 7). ΔC m увеличилась с 27,2 ± 4,7 фФ при комнатной температуре до 51,6 ± 6,0 фФ при высокой температуре ( p = 0,0006, n = 7).Q 10 для вызванного импульсом 20 мс ΔC m составляло 2,4 ± 0,4 ( n = 7).
Рисунок 8.
Эффективность экзоцитоза волосковых клеток усиливается при высокой температуре. A1 , B1 , ток Ca 2+ (I Ca ), емкость мембраны (C м ), сопротивление мембраны (R м ) и последовательное сопротивление (R ) s ) следы, полученные из волосковой клетки. Изменение ΔC м инициировалось ступенями деполяризации 20 мс ( A1 ) и 200 мс ( B1 ) от -90 мВ до -30 мВ при комнатной (черный) и высокой (красный) температуре.Для деполяризации 20 мс: я держу = -60 и -64 пА; C м (в состоянии покоя) = 14,5 и 16,0 пФ; R м = 1,3 и 1,2 ГОм; R s = 8,9 и 8,2 мОм при 24 ° C и 31 ° C соответственно. Для записей деполяризации 200 мс: I удерживает = -69 и -43 пА; C m (в состоянии покоя) = 11,3 и 12,8 пФ; R м = 1,2 и 1,5 ГОм; R s = 14,0 и 11,3 мОм при 24 ° C и 32 ° C соответственно. Эффективность экзоцитоза, рассчитанная как отношение ΔC m / Q Ca 2+ , показана для ( A2 ) 20 мс ( p = 0.0043, n = 7) и ( B2 ) 200 мс ( p = 0,0013, n = 15) деполяризации. Эффективность экзоцитоза почти в три раза выше для короткого импульса 20 мс, чем для длинного импульса 200 мс. ** p <0,01.
Здесь Q 10 — отношение скоростей реакции для T L = 296 ° K (23 ° C) и для T H = T L + 10 ° K = 306 ° K следующим образом :
где k B ≈ 8,3 Дж / моль ° K (постоянная Больцмана) и E a — энергетический барьер активации реакции.Из этого уравнения получаем E a = ( lnQ 10 ) · 75,2 кДж / моль. Таким образом, значение E a для вызванного экзоцитоза составило 65,8 ± 10,0 кДж / моль ≈ 26,8 k B T ( n = 7), что было рассчитано на основе Q 10 деполяризующих импульсов длительностью 20 мс. Это находится в пределах E и , оцененных для экзоцитоза везикул, опосредованного молекулами SNAREpin и синаптотагмина (20–140 k B T) (Zhang and Jackson, 2008; Schotten et al., 2015; Mostafavi et al., 2017), хотя экзоцитоз в волосковых клетках опосредуется отоферлином (Michalski et al., 2017).
Деполяризация от -90 до -30 мВ в течение 200 мс (Рис. 8 B1 ) не только истощает RRP, но также вызывает длительное высвобождение пузырьков, пополняющих RRP (Graydon et al., 2011). Q Ca , вызванный деполяризующим импульсом 200 мс, составлял 99,7 ± 8,0 пКл при 23,1 ± 0,2 ° C и 126,3 ± 9,7 пКл при 31,3 ± 0,2 ° C ( p <0,0001, n = 15).ΔC m , вызванная шагами деполяризации 200 мс, увеличилась с 109,8 ± 9,1 фФ при комнатной температуре до 192,1 ± 21,2 фФ при высокой температуре ( p <0,0001, n = 15). Q 10 Q Ca и ΔC m , вызванные импульсом 200 мс, составили 1,3 ± 0,04 и 2,0 ± 0,1 ( n = 15), соответственно. Q 10 экзоцитоза волосковых клеток лягушки близок к таковому, обнаруженному во внутренних волосковых клетках мыши с использованием измерений емкости (Q 10 = 2.1) (Nouvian, 2007) и в нейронах гиппокампа крысы с использованием методов обесцвечивания FM1-43 (Q 10 = 2,5) (Yang et al., 2005).
Построив график ΔC m относительно соответствующего Q Ca 2+ , мы получили эффективность экзоцитоза как отношение ΔC m / Q Ca 2+ , что для импульсов длительностью 20 мс увеличилось с 3,0 ± 0,8 фФ · пКл -1 при комнатной температуре до 4,7 ± 1,0 фФ · пКл -1 при высокой температуре ( p = 0.0043, n = 7; Q 10 = 1,9 ± 0,3; Рис.8 А2 ). Отношение ΔC м / Q Ca 2+ для откликов на импульсы длительностью 200 мс увеличилось с 1,2 ± 0,1 фФ · пКл −1 при комнатной температуре до 1,6 ± 0,2 фФ · пК −1 при высокой температуре ( p = 0,0013, n = 15; рис.8 B2 ). Записи ИГХ мышей также показывают, что более высокие температуры усиливают приток Ca 2+ и увеличивают ΔC m / Q Ca 2+ , вызванное деполяризующими импульсами длительностью 20 мс, но не для устойчивого экзоцитоза (Nouvian, 2007) .Напротив, наши результаты с волосковыми клетками лягушки-быка показывают, что эффективность как быстрого, так и устойчивого экзоцитоза увеличивается при высокой температуре.
Чтобы дополнительно продемонстрировать, что усилению экзоцитоза способствует не только увеличение притока Ca 2+ при высокой температуре, мы уменьшили приток Ca 2+ , используя меньшие деполяризующие потенциалы при высокой температуре, и измерили ΔC m скачков. Мы также использовали pH-нечувствительный буфер BAPTA Ca 2+ (Tsien, 1980) и другой pH-буфер MOPS, который имеет аналогичную температурную зависимость с HEPES (Ellis and Morrison, 1982).Деполяризация от -90 мВ до потенциала, который варьировался от -46 до -56 мВ (в среднем: -50,6 ± 1,8 мВ, n = 9) в течение 20 мс при 32 ° C, вызвала аналогичный или меньший приток Ca 2+ (6,4 ± 0,8 пКл) по сравнению с деполяризацией 20 мс до -30 мВ при 23 ° C (8,7 ± 1,2 пКл, p = 0,0059, n = 9; Рис. 9 A ). Однако даже когда приток Ca 2+ был уменьшен, волосковые клетки все еще были способны выделять больше пузырьков после деполяризации при высокой температуре (рис.9 А ). ΔC m увеличилась с 10,3 ± 1,3 фФ при комнатной температуре до 19,2 ± 2,6 фФ при высокой температуре ( p = 0,0028, n = 9; Рис.9 B ). В соответствии с предыдущими результатами, полученными при использовании внутреннего EGTA и HEPES, эффективность экзоцитоза увеличилась с 1,3 ± 0,2 при комнатной температуре до 3,7 ± 0,8 при высокой температуре ( p = 0,0042, n = 9; рис. 9 C ). Результаты показывают, что высокая температура способствует экзоцитозу синаптических везикул, даже когда приток Ca 2+ снижен.Более того, эффект не связан с температурно-зависимыми изменениями клеточного pH-зависимого буферизации Ca 2+ .
Рис. 9.
Экзоцитоз увеличивается при высокой температуре, даже когда приток Ca 2+ снижается. A , Потенциал деполяризации при высокой температуре регулировали между -45 и -55 мВ, чтобы вызвать такое же или меньшее количество притока Ca 2+ по сравнению с притоком при комнатной температуре. Чтобы исключить влияние температуры на pH или буферизацию Ca 2+ , волосковые клетки закрывали внутренним раствором, содержащим pH-нечувствительный буфер BAPTA для Ca 2+ (2 мм) и другой буфер pH MOPS (10 мм).Средние значения токов Ca 2+ (I Ca ) от девяти волосковых клеток, вызванных деполяризацией, до -30 мВ при 23 ° C (черный) и до -51 ± 2 мВ при 32 ° C (красный). I Ca был увеличен по шкале времени по сравнению с рамкой, изображенной пунктирной линией. Изменение емкости мембраны (ΔC м ) составило 10 фФ при 23 ° C (черный) и 19 фФ при 32 ° C (красный). B , ΔC m по сравнению с Q Ca из отдельных волосковых клеток деполяризованных до -30 мВ при 23 ° C (черный) и до -51 мВ при 32 ° C (красный).Точки данных, соединенные пунктирной линией, были записаны для одной и той же волосковой клетки. C , эффективность экзоцитоза (ΔC m / Q Ca 2+ ) увеличилась с 23 ° C (черный) до 32 ° C (красный; p = 0,0042, n = 9). ** p <0,01.
Размер RRP больше при высокой температуре
Чтобы сравнить максимальный размер RRP волосковой клетки при комнатной и высокой температуре, мы использовали протокол с двумя импульсами (Gillis et al., 1996). Выдаются два импульса длительностью 100 мс с интервалом 25 мс (рис. 10 A ). Короткий интервал 25 мс приводит к депрессии во втором выпуске. Ток Ca 2+ также может снижаться во время двойного импульса из-за инактивации. Чтобы избежать различий в Q Ca во время первой и второй деполяризации, потенциалы первых импульсов регулируются (рис. 10 A ). Среднее значение первых деполяризующих потенциалов составило -41,1 ± 0,7 мВ ( n = 11).Интервалы между парами импульсов составляли 60–90 с для полного восстановления. При температуре 23 ° C приток Ca 2+ во время первого и второго деполяризующих импульсов составил 31,8 ± 6,8 и 31,7 ± 6,8 пКл соответственно ( n = 4, p = 0,93; Рис.10 A , С ). При температуре 31 ° C приток Ca 2+ во время первого и второго деполяризующих импульсов составил 62,8 ± 3,8 и 63,3 ± 3,1 пКл соответственно ( n = 7, p = 0,69; Рис.10 A , С ).Соответственно, как первый ( p = 0,0019), так и второй ( p = 0,0009; рис. 10 C ) импульсы вызвали больший приток Ca 2+ при высокой температуре. ΔC 1 и ΔC 2 были скачками емкости, вызванными первой и второй деполяризацией. Максимальный размер RRP (B max ) был рассчитан согласно следующему:
Это уравнение от Gillis et al. (1996) основан на предположении, что одна и та же часть пула высвобождается с каждым импульсом.Если во время второго импульса наблюдается большая фракция высвобождения (например, из-за остаточного Ca 2+ из первого импульса) или значительное количество пузырьков пополнилось во время второго импульса, то B max будет завышать фактическое начальный размер пула. Следовательно, фактический размер пула находится между ΔC 1 и B max . В примере на Рисунке 10 A , B max увеличился со 143 пФ при 23 ° C до 175 пФ при 31 ° C. В среднем, B max увеличился с 83 ± 20 фФ ( n = 4) при комнатной температуре до 246 ± 47 фФ при высокой температуре ( n = 7, p = 0.0351, непарный т проба; Рис.10 B ). Эти результаты, показывающие увеличение размера RRP с повышением температуры, аналогичны тем, о которых сообщается для экзоцитоза хромаффинных клеток (Dinkelacker et al., 2000). Кроме того, отношение парных импульсов ΔC 2 / ΔC 1 существенно не изменилось при изменении температуры: 0,46 ± 0,19 при комнатной температуре ( n = 4) по сравнению с 0,69 ± 0,05 при высокой температуре ( n = 7, p = 0,18; непарный t тест).Это говорит о том, что вероятность вызванного выброса не изменяется при изменении температуры.
Рисунок 10.
RRP везикул больше при высокой температуре. A , Протокол парных импульсов, используемый для расчета максимального размера RRP, состоял из двух импульсов по 100 мс с интервалом 25 мс. Второй импульс был деполяризован до -30 мВ. Первый импульс был настроен так, чтобы обеспечить такое же общее количество притока Ca 2+ (Q Ca ), что и во втором импульсе. От камеры к камере все было по-разному.В этом примере первый деполяризующий потенциал составлял -38 мВ. Интервалы между парами импульсов составляли 60–90 с для полного восстановления. Ток Ca 2+ , зарегистрированный при 23 ° C и 31 ° C, показан черным и красным цветом соответственно. ΔC 1 и ΔC 2 — скачки емкости, вызванные первой и второй деполяризацией соответственно. B , Максимальный размер RRP был рассчитан согласно B max =. Значение B max значительно увеличилось с ∼83 fF ( n = 4) при 23 ° C до 246 fF при 31 ° C ( n = 7, p = 0.0351, непарный т проба). C , приток Ca 2+ во время первого и второго импульсов деполяризации составил 31,8 ± 6,8 пКл и 31,7 ± 6,8 пКл при комнатной температуре ( n = 4, p = 0,93), тогда как Ca 2+ приток во время первого и второго импульсов составил 62,8 ± 3,8 пКл и 63,3 ± 3,1 пКл при высокой температуре ( n = 7, p = 0,69). И первый ( p = 0,0005, непарный тест t ) и второй ( p = 0.0002, непарный t тест), таким образом, вызвали больший приток Ca 2+ при высокой температуре. * p <0,05, ** p <0,01, *** p <0,001.
Сверхбыстрые токи Ca
2+ и короткие синаптические задержки при высокой температуре
In vivo записи волокон слухового нерва двух разных видов лягушек, Hyla regilla и Eleutherodactylus coqui , показали, что латентность первый всплеск после появления звукового раздражителя уменьшался при повышении температуры с 12 ° C до 25 ° C (Stiebler and Narins, 1990).Чтобы определить температурную зависимость синаптической задержки в афферентном синапсе волосковых клеток, мы провели парные записи. Мы деполяризовали волосковые клетки с двумя шагами деполяризации по 20 мс от -90 до -30 мВ с интервалами 50 мс (рис. 11 A ) и зарегистрировали вызванные EPSC, опосредованные активацией рецепторов AMPA в афферентном волокне. После деполяризации ионы Ca 2+ вошли в волосковые клетки и запустили высвобождение глутамата (фиг. 11 A ). Из-за того, что EPSC сокращается в некоторых парных записях, мы включили в наш анализ только первое испытание из каждой пары.Таким образом, данные, полученные при комнатной и высоких температурах, относятся к двум разным группам пар. Среднее значение EPSC, зарегистрированных при 22 ° C ( n = 3) и 31 ° C ( n = 6), показано черным и красным цветом соответственно (рис. 11 A ). Синаптическая задержка измерялась от начала импульса до тех пор, пока EPSC не достигал 10% от своего максимума (рис. 11 C ). Синаптическая задержка первого EPSC (рис. 11 C ) уменьшилась с 2,03 ± 0,16 мс при комнатной температуре ( n = 3) до 1.28 ± 0,15 мс при высокой температуре ( n = 6, p = 0,0183, непарный тест t ; рис.11 E ). Синаптическая задержка второго EPSC (рис.11 C ) сократилась с 1,26 ± 0,10 мс при комнатной температуре ( n = 3) до 0,72 ± 0,05 мс при высокой температуре ( n = 6, p = 0,001, непарный t тест; рис.11 E ). Первая синаптическая задержка была больше второй под обеими комнатами ( n = 3, p = 0.008) и высокой ( n = 6, p = 0,0206) температурой (рис.11 D , E ). Синаптическая задержка, которую мы измерили по синапсам волосковых клеток взрослых лягушек, намного короче, чем задержка синапсов внутренних волосковых клеток незрелых крыс (Goutman and Glowatzki, 2011).
Рисунок 11.
Более короткие синаптические задержки и более быстрые EPSC при высокой температуре. A , Два шага деполяризации по 20 мс от -90 мВ до -30 мВ с интервалами 50 мс были применены к волосковой клетке. Одновременно постсинаптическое афферентное волокно было зафиксировано напряжением -90 мВ.Среднее значение притока кальция (I Ca ), вызванного стимулом волосковых клеток при 22 ° C ( n = 3) и 31 ° C ( n = 6) показано черным и красным, соответственно. Высвобождение глутамата из синаптических пузырьков из волосковой клетки запускает многоквантовый EPSC в афферентном волокне. Черные и красные кривые представляют собой усредненные значения EPSC при 22 ° C ( n = 3) и 31 ° C ( n = 6) соответственно. Из-за значительного сокращения мы проанализировали только первое испытание каждой пары. Таким образом, ответы I Ca и EPSC при комнатной и высокой температуре были получены от двух разных групп пар. B , Представитель Ca 2+ токи от другой волосковой клетки были вызваны деполяризацией 20 мс от -90 мВ до -30 мВ при 24 ° C (черный) и 32 ° C (красный) с использованием P / 4 протокол для вычитания утечек. Постоянная времени активации была получена из однократной экспоненциальной подгонки после емкостного переходного процесса (пунктирные линии). В этом примере постоянные времени активации составляли 366 мкс при 24 ° C (черный) и 204 мкс при 32 ° C (красный). C , Фаза нарастания первого EPSC показана в расширенном масштабе времени.Синаптическая задержка определялась как время между началом стимуляции и достижением EPSC 10% от максимума. D , Наложение первого (пунктирные линии) и второго (сплошные линии) EPSC. Обратите внимание на большое синаптическое облегчение вторых EPSC. E . Синаптическая задержка как первого ( p = 0,0183, непарный тест t ), так и второго EPSC ( p = 0,001, непарный тест t ) уменьшалась при 31 ° C.Первая синаптическая задержка была больше второй как при 22 ° C ( n = 3, p = 0,008), так и при 31 ° C ( n = 6, p = 0,02). * p <0,05, ** p <0,01.
Более короткая синаптическая задержка частично связана с более быстрой активацией тока Ca 2+ при высокой температуре. Чтобы определить влияние температуры на активацию I Ca , мы использовали вычитание утечки P / 4 для устранения емкостного переходного процесса.Ток Ca 2+ был вызван шагом деполяризации 20 мс от -90 до -30 мВ (рис. 11 B ). Подгоняя фазу активации тока Ca 2+ одной экспоненциальной функцией (рис.11 B , пунктирные линии), мы вычислили постоянную времени активации: 377 ± 30 мкс при 23,9 ± 0,1 ° C и 219 ± 19 мкс при 31,5 ± 0,3 ° C ( p <0,0001, n = 8). Соответствующий Q 10 составил 2,2 ± 0,2 ( n = 8), что аналогично Q 10 активации каналов Ca 2+ L-типа в сенсорных нейронах цыплят (Acerbo and Nobile, 1994), но выше, чем Q 10 внутренних волосковых клеток песчанок (Johnson and Marcotti, 2008).Результаты показывают, что высокая температура ускоряет активацию каналов Ca 2+ и, таким образом, способствует сокращению синаптической задержки.
Коэффициент парных импульсов, рассчитанный из переноса заряда EPSC (EPSC 2 / EPSC 1 ), составил 1,00 ± 0,05 при комнатной температуре ( n = 3) и 0,92 ± 0,02 при высокой температуре ( n = 6, p = 0,25, непарный t тест). Вместе с коэффициентом парных импульсов ΔC 2 / ΔC 1 это говорит о том, что высокая температура не влияет на общую вероятность выброса.Однако высокая температура улучшает временную точность за счет сокращения синаптической задержки; и при обеих температурах синапсы волосковых клеток обнаруживают выраженное кратковременное облегчение (Goutman and Glowatzki, 2011; см. также Cho et al., 2011). Интересно, что эти результаты сильно отличаются от результатов, обнаруженных в обычном синапсе активной зоны, чашечке синапса Held, где синаптическая задержка второго импульса была больше, а EPSC обнаруживают выраженную кратковременную депрессию (Wu and Borst, 1999).
Обсуждение
Температурные изменения EPSC, EPSP и скорости спайков
Наши записи in vitro целых клеток афферентных волокон показывают, что частоты EPSC и EPSP увеличиваются при более высоких температурах.Это должно привести к увеличению всплесков AP при более высоких температурах. Действительно, частота спайков афферентных волокон в основном увеличивается при более высоких температурах (рис. 1). In vivo записи одиночных волокон слухового нерва также показывают, что частота спонтанных спайков в волокнах сосочка земноводных лягушки-быка увеличивается при более высоких температурах тела (van Dijk et al., 1990). Однако около 30% наших записей афферентных волокон снижали скорость их спайков при более высоких температурах. Мы не знаем причин этой неоднородности.Основные механизмы, которые запускают спайки в различных афферентных волокнах, требуют дополнительных исследований, чтобы объяснить широкий диапазон входного сопротивления мембраны (Рис. 4 B ). Интересно, что волокна слухового нерва млекопитающих также демонстрируют неоднородность по частоте спайков, экспрессии генов и свойствам мембран (Rutherford et al., 2012; Heil and Peterson, 2017; Shrestha et al., 2018).
Какие механизмы вызывают увеличение частоты EPSC? Одна очевидная возможность, которую мы исследовали, была деполяризация волосковой клетки V rest .Неожиданно оказалось, что во время записи токовым зажимом целых клеток, V оставил волосковых клеток, значительно гиперполяризованных (фиг. 5 A ). V rest также гиперполяризуется в нейронах улитки при более высоких температурах из-за повышенной активности Na + / K + -АТФазы (Gorman and Marmor, 1970). Однако записи перфорированных пластырей, которые не нарушают внутриклеточную среду, не показали значительных изменений. Остаток V , который мы наблюдали в режиме перфорированного пятна, составлял ∼ -63 мВ (рис.5 А ). Предыдущие четкие записи обычных электродов V , оставшиеся волосковых клеток черепахи и лягушки, показали значения ~ -55 мВ (Crawford and Fettiplace, 1980; Pitchford and Ashmore, 1987). При этих мембранных потенциалах индуцируется небольшой входящий внутрь ток Ca 2+ (рис. 5 C ). Этот ток покоя Ca 2+ увеличивается при более высоких температурах, что, вероятно, увеличивает вероятность высвобождения и скорость всплесков (Trapani and Nicolson, 2011). Частота спонтанных mEPSC также увеличивается при высокой температуре в чашечке синапса Held (Kushmerick et al., 2006; Postlethwaite et al., 2007) и синапсы гиппокампа (Kim and Connors, 2012). Однако, в отличие от волосковых клеток лягушки-быка, V покоится на чашечках нервных окончаний Held деполяризуется при повышении температуры с 24 ° C до 32 ° C (Kim and von Gersdorff, 2012). Наконец, мы отмечаем, что наши волосковые клетки сосочка амфибий купаются в высоком внешнем Ca 2+ , который блокирует каналы трансдукции; и недавно было обнаружено, что внезапное повышение температуры может смягчить запирающие пружины и открыть каналы механотрансдукции на пучках стереоцилий волосковых клеток саккулюса лягушки (Azimzadeh et al., 2018). Следовательно, мы можем недооценивать физиологические эффекты температурных сдвигов на волосковую клетку, и V rest может быть более деполяризованным в условиях in vivo .
Мы обнаружили, что средняя амплитуда спонтанных ЭПСК от афферентных волокон лягушки не изменилась при более высокой температуре (рис. 2; табл. 2). Это отличается от наблюдений, сделанных в синапсах млекопитающих, где повышение температуры увеличивает спонтанную амплитуду mEPSC (Veruki et al., 2003; Кушмерик и др., 2006; Postlethwaite et al., 2007). Интересно, что ранее мы наблюдали, что снижение температуры от комнатной до 15 ° C привело к уменьшению амплитуды EPSC (Li et al., 2009). Таким образом, спонтанные амплитуды EPSC могут быть насыщенными при комнатной температуре. Средняя амплитуда ВПСП также не менялась с температурой (табл. 2). Однако в 13 волокнах, где были зарегистрированы как EPSC, так и EPSP, EPSC остались прежними ( p = 0,45), тогда как амплитуда EPSP уменьшилась ( p = 0.049, n = 13) при высокой температуре. Это могло быть результатом уменьшения афферентного волокна R в при высокой температуре. Частота событий EPSC и EPSP увеличивалась при высокой температуре (Таблица 2). Это может привести к большему суммированию EPSP, повышая вероятность того, что они достигнут порога AP. Однако распад ВПСП также был значительно быстрее при более высоких температурах (рис. 2 A3 , B3 ), что имеет тенденцию уменьшать суммирование несовпадающих ВПСП. Следовательно, возможно, что более высокие температуры могут снизить частоту всплесков, как показано на Рисунке 1 B .Таким образом, как пресинаптические, так и постсинаптические механизмы могут объяснить неоднородность изменений частоты спайков в зависимости от температуры, показанную на Рисунке 1.
Температурные изменения порогового значения спайков также могут объяснить неоднородность, показанную на Рисунке 1. Термоположительные волокна спонтанно генерируют больше спайков на высоких уровнях. температура с уменьшением порога на ∼5 мВ, тогда как термоотрицательные волокна генерируют меньше спонтанных всплесков при высокой температуре с увеличением порога на 5 мВ (Таблица 1). Неоднородность типов и плотностей потенциалзависимых каналов Na + и K + среди разных волокон может объяснить эти различия (Kim and Rutherford, 2016).В совокупности температурно-зависимые изменения скорости спайков обусловлены пресинаптическими изменениями экзоцитоза волосковых клеток и постсинаптическими изменениями возбудимости афферентных волокон.
Синаптическая задержка и фазовая синхронизация при высокой температуре
Записи одиночных волокон слухового нерва лягушки показывают, что задержка начала спайков на звуковой стимул уменьшалась с увеличением температуры со значениями Q 10 1,6–2,5 (Stiebler and Narins, 1990; van Dijk и др., 1990). Мы выполнили парные записи от волосковой клетки до афферентного волокна и выявили сокращение синаптической задержки при высокой температуре (рис.11 E ). Эта более короткая синаптическая задержка частично объясняется более быстрой кинетикой активации тока Ca 2+ при высокой температуре (Рис. 11 B ). Кроме того, постоянная времени слияния липидных мембран обратно коррелирует с температурой (Schotten et al., 2015), что также может вносить вклад в более короткую синаптическую задержку при высокой температуре. Более короткая синаптическая задержка улучшит синхронизацию фазы спайков волокон, потому что вызванные ВПСП и спайки будут в большей степени синхронизированы по фазе с текущей активацией волосковых клеток Ca 2+ .Это может объяснить, почему in vivo записи отдельных волокон слухового нерва указывают на то, что сила переносчика увеличивается при более высоких температурах (Stiebler and Narins, 1990; van Dijk et al., 1990).
Более высокие температуры и синаптическое облегчение имеют тенденцию синхронизировать вызванное деполяризацией мультивезикулярное высвобождение (Fig. 11). Синаптическое облегчение, выявленное деполяризацией парных импульсов, также снижает синаптические задержки как в синапсах волосковых клеток лягушки (Cho et al., 2011), так и в синапсах внутренних волосковых клеток крыс (синаптическая задержка равна 1.5 мс при комнатной температуре) (Goutman, Glowatzki, 2011). Аналогичным образом, мы измерили здесь синаптические задержки 1,3 мс при 23 ° C и 0,7 мс при 31 ° C (рис. 11 E ).
Электрический резонанс при высокой температуре
Снижение R в при высокой температуре уменьшает постоянную времени мембраны, позволяя волосковым клеткам лучше отслеживать быстрые изменения мембранного потенциала (рис. 6). Собственная электрическая резонансная частота волосковых клеток увеличивается с температурой (рис.7), как и ожидалось из-за увеличения амплитуд тока Ca 2+ и K + (рис.5 В ). Мы наблюдали большое изменение средней резонансной частоты от 320 до 530 Гц (рис. 7 D ). Эти изменения увеличивают динамический диапазон частотной перестройки волосковых клеток сосочка земноводных и связанных с ними афферентных волокон. Действительно, in vivo записи волокон слухового нерва лягушки показывают более высокие характеристические частоты (вершина кривой настройки или лучшая частота) при более высокой температуре (Stiebler and Narins, 1990).
Эффективность экзоцитоза и размер пула везикул при высокой температуре
Меньший R в неизбежно приведет к меньшим изменениям мембранного потенциала (V m = R в × I), вызванным открытием механочувствительных каналов в стереоцилиях.Такой прогнозируемой потери чувствительности к звуковым сигналам не наблюдается in vivo . В действительности наблюдается обратное: порог спайков, вызванных звуком, снижается при высокой температуре (Stiebler and Narins, 1990). Как мы можем объяснить это повышение чувствительности волокна?
Во-первых, мы обнаружили, что крутизна кривой тока Ca 2+ I – V (рис. 5 C ) увеличивается при высокой температуре, что делает приток Ca 2+ более чувствительным к колебаниям мембранного потенциала около V остальное .Увеличенный ток Ca 2+ может быть результатом увеличения вероятности открытия канала Ca 2+ L-типа (P o ) и / или проводимости (Lux and Brown, 1984; Klöckner et al., 1990; Acerbo и Nobile, 1994; Peloquin et al., 2008). Q 10 проводимости одного канала L-типа в гладкомышечных клетках и сетчатке составляет 1,6 и 3,0, соответственно (Klöckner et al., 1990; Peloquin et al., 2008). Во-вторых, в синапсах волосковых клеток кластер каналов Ca 2+ L-типа расположен в пределах нанометров от пристыкованных пузырьков под синаптической лентой (Brandt et al., 2005; Nouvian et al., 2006; Graydon et al., 2011; Ким и др., 2013). Быстрая активация этого кластера каналов Ca 2+ дает высокосинхронный и быстрый многоквантовый EPSC (рис.11 D ) (Grabner et al., 2016), что может помочь объяснить короткую задержку и низкий уровень джиттера звук вызвал первый спайк в слуховом нерве (Wittig and Parsons, 2008).
Мы также наблюдали увеличение эффективности экзоцитоза, что указывает на то, что увеличение экзоцитоза не пропорционально увеличению тока Ca 2+ .Нелинейная степенная зависимость экзоцитоза от концентрации свободных ионов Ca 2+ может потенциально объяснить наши результаты высокого Q 10 = 2,4 для экзоцитоза, вызванного деполяризующими импульсами длительностью 20 мс (Cho and von Gersdorff, 2012; Quiñones et al. , 2012). Однако линейная зависимость экзоцитоза от заряда тока Ca 2+ была обнаружена для среднечастотных настроенных волосковых клеток лягушки быка (Keen and Hudspeth, 2006; Cho et al., 2011). Таким образом, эффективность слияния везикул должна быть существенно выше при высокой температуре, как показано на Рисунке 9 C .В дополнение к притоку Ca 2+ через каналы Ca 2+ L-типа в плазматической мембране, вызванное Ca 2+ высвобождение Ca 2+ из внутриклеточных хранилищ также может участвовать в нелинейном рекрутировании пузырьков в слуховой проход. волосковые клетки (Castellano-Muñoz et al., 2016).
Вероятность выброса увеличивается при высокой температуре для некоторых центральных синапсов (Hardingham, Larkman, 1998; Волгушев и др., 2004), хотя мы не наблюдали изменений в вероятности выброса при использовании протокола стимуляции парными импульсами (рис.10, 11). Однако мы действительно наблюдали значительное увеличение размера RRP при высокой температуре (рис. 10). Помимо облегчения спонтанного слияния пузырьков, температура также способствует устойчивому экзоцитозу за счет ускорения пополнения пузырьков RRP (Pyott and Rosenmund, 2002; Kushmerick et al., 2006). Более того, эндоцитоз синаптических пузырьков также очень чувствителен к температуре (Fernández-Alfonso and Ryan, 2004; Micheva and Smith, 2005; Renden and von Gersdorff, 2007; Delvendahl et al., 2016).Таким образом, кинетика эндоцитоза может ограничивать скорость длительного экзоцитоза и требует дальнейших исследований синапсов волосковых клеток.
7.3 Череп — анатомия и физиология
Цели обучения
К концу этого раздела вы сможете:
- Перечислить и идентифицировать кости черепа и лицевого черепа и определить их важные особенности
- Найдите основные линии швов черепа и назовите сочленяющиеся кости, образующие их
- Определите придаточные пазухи носа и определите расположение каждой
- Назовите кости, которые составляют стенки орбиты, и определите отверстия, связанные с орбитой
- Определите кости и структуры, которые образуют носовую перегородку и носовые раковины, и найдите подъязычную кость
- Определить костные отверстия черепа
Череп — это скелетная структура головы, которая поддерживает лицо и защищает мозг.Он подразделяется на лицевых костей и черепа или свода черепа (рис. 7.3.1). Лицевые кости лежат в основе структур лица, образуют носовую полость, охватывают глазные яблоки и поддерживают зубы верхней и нижней челюстей. Округлый череп окружает и защищает мозг, в нем находятся структуры среднего и внутреннего уха.
У взрослого человека череп состоит из 22 отдельных костей, 21 из которых неподвижны и объединены в единое целое. 22-я кость — это нижняя челюсть (нижняя челюсть), которая является единственной подвижной костью черепа.
Рисунок 7.3.1 — Части черепа: Череп состоит из округлой формы черепа, в которой находится мозг, и лицевых костей, образующих верхнюю и нижнюю челюсти, нос, орбиты и другие лицевые структуры.
Внешний веб-сайт
Посмотрите это видео, чтобы увидеть вращающийся взорванный череп с разноцветными костями. Какая кость (желтая) расположена в центре и соединяется с большинством других костей черепа?
Передняя часть черепа состоит из лицевых костей и обеспечивает костную опору для глаз, зубов и структур лица, а также обеспечивает отверстия для еды и дыхания.В этом виде черепа преобладают отверстия глазниц и носовой полости. Также видны верхняя и нижняя челюсти с соответствующими зубами (рис. 7.3.2).
Орбита — это костная впадина, в которой находится глазное яблоко и мышцы, которые двигают глазное яблоко или открывают верхнее веко. Верхний край передней орбиты — надглазничный край . Рядом с серединой надглазничного края расположено небольшое отверстие, называемое надглазничным отверстием .Это обеспечивает прохождение чувствительного нерва к коже лба. Ниже орбиты находится подглазничное отверстие , которое является точкой выхода сенсорного нерва, который снабжает переднюю часть лица ниже орбиты.
Рисунок 7.3.2 — Вид черепа спереди: На виде черепа спереди показаны кости, которые образуют лоб, орбиты (глазницы), полость носа, носовую перегородку, а также верхнюю и нижнюю челюсти.
Внутри носовой области черепа носовая полость разделена пополам носовой перегородкой .Верхняя часть носовой перегородки образована перпендикулярной пластиной решетчатой кости , а нижняя часть — сошниковой костью . Если смотреть в носовую полость спереди черепа, видны две костные пластинки, выступающие из каждой боковой стенки. Более крупная из них — нижняя носовая раковина , независимая кость черепа. Чуть выше нижней раковины расположена средняя носовая раковина , которая является частью решетчатой кости.Третья костная пластинка, также являющаяся частью решетчатой кости, — это верхняя носовая раковина . Он намного меньше по размеру и находится вне поля зрения, над средней раковиной. Верхняя носовая раковина расположена латеральнее перпендикулярной пластинки в верхней носовой полости.
На виде бокового черепа преобладает большой округлый череп сверху, а также верхняя и нижняя челюсти с зубами снизу (рис. 7.3.3). Эти области разделяет костный мост, называемый скуловой дугой. скуловая дуга (скула) — это костная дуга на боковой стороне черепа, которая простирается от области щеки до чуть выше слухового прохода.Он образован стыком двух костных отростков: короткого переднего компонента, височного отростка скуловой кости и более длинной задней части, скулового отростка височной кости , идущего вперед от височной кости. Таким образом, височный отросток (спереди) и скуловой отросток (сзади) соединяются вместе, как два конца подъемного моста, образуя скуловую дугу. Одна из основных мышц, которая тянет нижнюю челюсть вверх во время кусания и жевания, жевательная мышца, исходит из скуловой дуги.
На боковой стороне черепа, выше уровня скуловой дуги, находится неглубокое пространство, называемое височной ямкой . Возникающая из височной ямки и проходящая глубоко в скуловую дугу — это еще одна мышца, которая действует на нижнюю челюсть во время жевания, — височную.
Рисунок 7.3.3 — Боковой вид и сагиттальный разрез черепа: (a) Боковой вид черепа. На боковой стороне черепа виден большой округлый корпус мозга, скуловая дуга, а также верхняя и нижняя челюсти.Скуловая дуга образована совместно скуловым отростком височной кости и височным отростком скуловой кости. Неглубокое пространство над скуловой дугой — это височная ямка. (б) Сагиттальный разрез черепа. На этом изображении средней линии черепа в сагиттальном разрезе показана носовая перегородка.
Череп содержит и защищает мозг. Внутреннее пространство, которое почти полностью занято мозгом, называется полостью черепа . Эта полость ограничена сверху закругленной вершиной черепа, которая называется сводом черепа, (черепная крышка), а также боковой и задней сторонами черепа.Кости, образующие верхнюю и боковые части черепа, обычно называют «плоскими» костями черепа.
Пол корпуса головного мозга называется основанием черепа или дном черепа. Это сложная область, которая различается по глубине и имеет множество отверстий для прохождения черепных нервов, кровеносных сосудов и спинного мозга. Внутри черепа основание разделено на три больших пространства, называемых передней черепной ямкой , средней черепной ямкой и задней черепной ямкой (ямка = «траншея или канавка») (Рисунок 7.3.4). Глубина ямок увеличивается от переднего к заднему. Форма и глубина каждой ямки соответствуют форме и размеру области мозга, в которой находится каждая из них.
Рисунок 7.3.4 — Черепные ямки: Кости корпуса мозга окружают и защищают мозг, который занимает полость черепа. Основание корпуса мозга, которое образует дно полости черепа, подразделяется на мелкую переднюю черепную ямку, среднюю черепную ямку и глубокую заднюю черепную ямку.
Череп состоит из восьми костей. К ним относятся парные теменные и височные кости, а также непарные лобные, затылочные, клиновидные и решетчатые кости.
Теменная кость
теменная кость образует большую часть верхней и боковой стороны черепа (см. Рисунок 7.3.3). Это парные кости, при этом правая и левая теменные кости соединяются вместе в верхней части черепа, образуя сагиттальный шов. Каждая теменная кость также ограничена спереди лобной костью у коронарного шва, снизу височной костью у плоского шва и сзади затылочной костью у лямбдовидного шва.
Рисунок 7.3.5 . Улучшенный вид черепа.
височная кость
височная кость образует нижнюю боковую сторону черепа (см. Рисунок 7.3.3). Распространенная мудрость гласит, что височная кость (temporal = «время») названа так, потому что именно в этой области головы (виске) волосы обычно сначала становятся седыми, указывая на течение времени.
Височная кость подразделяется на несколько областей (рис. 7.3.6). Уплощенная верхняя часть — это плоская часть височной кости.Ниже этой области выступает вперед скуловой отросток височной кости, который образует заднюю часть скуловой дуги. Кзади находится сосцевидная часть височной кости. Внизу от этой области выступает большой выступ, сосцевидный отросток , который служит местом прикрепления мышц. Сосцевидный отросток легко прощупывается на стороне головы сразу за мочкой уха. На внутренней стороне черепа каменистая часть каждой височной кости образует выступающий вертикальный, диагонально ориентированный каменистый гребень , который поднимается от задней черепной ямки к средней черепной ямке.Внутри каждого каменистого гребня расположены небольшие полости, в которых находятся структуры среднего и внутреннего ушей.
Рисунок 7.3.6 — Височная кость: На виде сбоку изолированной височной кости показаны плоскоклеточная, сосцевидная и скуловая части височной кости.
Важные ориентиры височной кости, как показано на рисунке 7.3.7, включают следующее:
- Внешний слуховой проход (слуховой проход) — это большое отверстие на боковой стороне черепа, которое связано с ухом.
- Внутренний слуховой проход — Это отверстие находится внутри полости черепа, на медиальной стороне каменистого гребня. Он соединяется с полостями среднего и внутреннего уха височной кости.
- Нижнечелюстная ямка — Это глубокое углубление овальной формы, расположенное на внешнем основании черепа, прямо перед наружным слуховым проходом. Нижняя челюсть (нижняя челюсть) соединяется с черепом в этом месте как часть височно-нижнечелюстного сустава, что позволяет выполнять движения нижней челюсти во время открывания и закрывания рта.
- Суставной бугорок — Гладкий гребень, расположенный непосредственно перед нижнечелюстной ямкой. И суставной бугорок, и нижнечелюстная ямка участвуют в работе височно-нижнечелюстного сустава, сустава, обеспечивающего движения между височной костью черепа и нижней челюстью.
- Шиловидный отросток — Сзади от нижней челюстной ямки на внешнем основании черепа находится удлиненный, направленный вниз костный выступ, называемый шиловидным отростком, названный так из-за его сходства со стилусом (ручкой или пишущим инструментом).Эта структура служит местом прикрепления нескольких мелких мышц и связки, поддерживающей подъязычную кость шеи. (См. Также рисунок 7.36.)
- Сосцевидное отверстие — Это небольшое отверстие расположено между шилососцевидным отростком и сосцевидным отростком. Это точка выхода черепного нерва, который питает лицевые мышцы.
- Каротидный канал — Каротидный канал представляет собой зигзагообразный туннель, который обеспечивает проход через основание черепа для одной из основных артерий, кровоснабжающих мозг.Его вход расположен на внешней стороне основания черепа, перед шиловидным отростком и непосредственно перед яремным отверстием. Затем канал проходит в переднемедиальном направлении внутри костной основы черепа, а затем поворачивает вверх к своему выходу в дне средней черепной полости, над кружевным отверстием.
- Яремное отверстие — Отверстие в височной кости непосредственно позади сонного канала. Это точка выхода внутренней яремной вены.
Рисунок 7.3.7 — Внешний и внутренний вид основания черепа: (a) Твердое небо образовано спереди небными отростками верхнечелюстных костей, а сзади — горизонтальной пластиной небных костей. (б) Сложное дно полости черепа образовано лобной, решетчатой, клиновидной, височной и затылочной костями. Малое крыло клиновидной кости разделяет переднюю и среднюю черепные ямки. Каменистый гребень (каменистая часть височной кости) разделяет среднюю и заднюю черепные ямки.
Лобная кость
лобная кость — единственная кость, которая образует лоб. На его передней средней линии, между бровями, имеется небольшое углубление, называемое глабелью (см. Рис. 7.3.3). Лобная кость также образует надглазничный край глазницы. Ближе к середине этого края находится надглазничное отверстие, отверстие, через которое проходит сенсорный нерв ко лбу. Лобная кость утолщена чуть выше каждого надглазничного края, образуя округлые надбровные дуги.Они расположены сразу за бровями и различаются по размеру у разных людей, хотя, как правило, у мужчин они больше. Внутри полости черепа кзади выходит лобная кость. Эта уплощенная область образует как крышу орбиты внизу, так и дно передней полости черепа вверху (см. Рис. 7.3.7 b ).
Затылочная кость
Затылочная кость — единственная кость, которая образует задний череп и заднюю черепную ямку (Рисунок 7.3,8; см. также рисунок 7.3.7). На его внешней поверхности, на задней средней линии, есть небольшой выступ, называемый внешним затылочным выступом , который служит местом прикрепления связки задней части шеи. Латерально по обе стороны от этой выпуклости проходит верхняя линия шеи (выйная зона = «затылок» или «задняя часть шеи»). Линии шеи представляют собой самую верхнюю точку, в которой мышцы шеи прикрепляются к черепу, при этом только скальп покрывает череп выше этих линий.В основании черепа затылочная кость содержит большое отверстие большого затылочного отверстия , через которое спинной мозг выходит из черепа. По обе стороны от большого затылочного отверстия находится затылочный мыщелок овальной формы . Эти мыщелки образуют суставы с первым шейным позвонком, которые позволяют кивать (как согласованное) движение головы.
Рисунок 7.3.8 — Задний вид черепа: На этом виде задней части черепа показаны места прикрепления мышц и суставов, которые поддерживают череп.
Клиновидная кость
Клиновидная кость — это отдельная сложная кость центрального черепа (рис. 7.3.9). Она служит «краеугольным камнем», потому что соединяется почти со всеми остальными костями черепа. Клиновидная кость образует большую часть основания центрального черепа (см. Рис. 7.3.7), а также расширяется в боковом направлении, внося вклад в боковые стороны черепа (см. Рис. 7.3.3). Внутри полости черепа правое и левое малых крыльев клиновидной кости , которые напоминают крылья летящей птицы, образуют выступ выступающего гребня, который отмечает границу между передней и средней черепными ямками. sella turcica («турецкое седло») расположено по средней линии средней черепной ямки. Эта костная область клиновидной кости названа из-за сходства с конскими седлами, использовавшимися турками-османами, с высокой спинкой, называемой dorsum sellae , и высокой передней частью. Округленное углубление на дне турецкого седла — это гипофизарная (гипофизарная) ямка , в которой находится гипофиз (гипофиз) размером с горошину. большие крылья клиновидной кости проходят латерально в обе стороны от турецкого седла, где они образуют переднее дно средней черепной ямки.Большое крыло лучше всего видно на внешней стороне бокового черепа, где оно образует прямоугольную область непосредственно перед плоскоклеточной частью височной кости.
На нижней стороне черепа каждая половина клиновидной кости образует две тонкие вертикально ориентированные костные пластинки. Это медиальная крыловидная пластина и латеральная крыловидная пластина (крыловидная пластина = «крыловидная»). Правая и левая медиальные крыловидные пластинки образуют заднюю, боковую стенки носовой полости.Несколько более крупные латеральные крыловидные пластинки служат местами прикрепления жевательных мышц, заполняющих подвисочное пространство и действующих на нижнюю челюсть.
Важные ориентиры клиновидной кости, как показано на рисунке 7.3.9.
- Зрительный канал — Это отверстие расположено в переднем боковом углу турецкого седла. Он обеспечивает выход зрительного нерва в орбиту.
- Верхняя глазничная щель — Это большое отверстие неправильной формы, ведущее в заднюю орбиту, расположено на передней стенке средней черепной ямки, латеральнее зрительного канала и под выступающим краем малого крыла клиновидной кости.Через это отверстие проходят нервы глазного яблока и связанных с ним мышц, а также сенсорные нервы лба.
- Foramen rotundum — Это закругленное отверстие (rotundum = «круглый») расположено в дне средней черепной ямки, чуть ниже верхней глазничной щели. Это точка выхода главного сенсорного нерва, который питает щеку, нос и верхние зубы.
- Овальное отверстие средней черепной ямки — Это большое овальное отверстие в дне средней черепной ямки обеспечивает проход для основного сенсорного нерва к боковой головке, щеке, подбородку и нижним зубам.
- Foramen spinosum — Это небольшое отверстие, расположенное кзади-латеральнее овального отверстия, является точкой входа для важной артерии, которая снабжает покровные слои, окружающие мозг. Схема ветвления этой артерии образует хорошо заметные бороздки на внутренней поверхности черепа, и эти борозды можно проследить до их происхождения в остистом затылке.
- Каротидный канал — Это зигзагообразный проход, через который в череп входит основная артерия, ведущая к головному мозгу.Вход в сонный канал расположен в нижней части черепа, перед шиловидным отростком (см. Рис. 7.3.7 a ). Отсюда канал проходит переднемедиально в костном основании черепа. Сразу над кружевным отверстием сонный канал открывается в среднюю полость черепа, рядом с заднебоковым основанием турецкого седла.
- Foramen lacerum — Это отверстие неправильной формы расположено в основании черепа, непосредственно ниже выхода сонного канала.Это отверстие — артефакт сухого черепа, потому что при жизни он полностью заполнен хрящом. Все отверстия черепа, через которые проходят нервы или кровеносные сосуды, имеют гладкие края; слово Lacerum («рваный» или «разорванный») говорит нам, что это отверстие имеет неровные края, и поэтому через него ничего не проходит.
Рисунок 7.3.9 — Клиновидная кость: Клиновидная кость, изображенная изолированно на (а) верхнем и (б) заднем изображениях, представляет собой единственную срединную кость, которая образует передние стенки и дно средней черепной ямки.У него есть пара меньших крыльев и пара больших крыльев. Турецкое седло окружает гипофизарную ямку. Снизу выступают медиальная и латеральная крыловидные пластинки. Клиновидная кость имеет несколько отверстий для прохождения нервов и кровеносных сосудов, включая зрительный канал, верхнюю глазничную щель, круглое отверстие, овальное и остистое отверстие.
Решетчатая кость
решетчатая кость — это единственная кость по средней линии, которая образует крышу и боковые стенки верхней носовой полости, верхней части носовой перегородки и способствует медиальной стенке глазницы (Рисунок 7.3.10 и рисунок 7.3.11). На внутренней стороне черепа решетчатая кость также образует часть дна передней полости черепа (см. Рис. 7.3.7 b ).
Внутри полости носа перпендикулярная пластинка решетчатой кости образует верхнюю часть носовой перегородки. Решетчатая кость также образует боковые стенки верхней носовой полости. От каждой боковой стенки отходят верхняя носовая раковина и средняя носовая раковина, которые представляют собой тонкие изогнутые выступы (носовые раковины), которые заходят в носовую полость (Рисунок 7.3.12).
В полости черепа решетчатая кость образует небольшой участок по средней линии дна передней черепной ямки. Эта область также образует узкую крышу подлежащей носовой полости. Эта часть решетчатой кости состоит из двух частей: crista galli и ребристых пластин. crista galli («гребешок или гребень петуха») представляет собой небольшой костный выступ, направленный вверх, расположенный по средней линии. Он функционирует как передняя точка прикрепления одной из мозговых оболочек (защитных мембран, покрывающих головной мозг).По обе стороны от crista galli находится пластинка cribriform (cribrum = «сито»), небольшая уплощенная область с многочисленными небольшими отверстиями, называемыми обонятельными отверстиями. Маленькие нервные ветви из обонятельных областей носовой полости проходят через эти отверстия и попадают в мозг.
Боковые части решетчатой кости расположены между глазницей и верхней полостью носа и, таким образом, образуют боковую стенку полости носа и часть медиальной стенки глазницы. Внутри этой части решетчатой кости расположены несколько небольших заполненных воздухом пространств, которые являются частью системы придаточных пазух носа черепа.
Рисунок 7.3.10 — Решетчатая кость: Непарная решетчатая кость расположена по средней линии внутри центрального черепа. Он имеет выступ вверх, crista galli, и выступ вниз, перпендикулярную пластинку, которая образует верхнюю перегородку носа. Решетчатые пластинки образуют как крышу носовой полости, так и часть дна передней черепной ямки. Боковые стороны решетчатой кости образуют боковые стенки верхней носовой полости, часть медиальной стенки глазницы и дают начало верхней и средней носовым раковинам.Решетчатая кость также содержит решетчатые воздушные клетки.
Рисунок 7.3.11 — Боковая стенка носовой полости: Три носовых раковины представляют собой изогнутые кости, которые выступают из боковых стенок носовой полости. Верхняя носовая раковина и средняя носовая раковина являются частями решетчатой кости. Нижняя носовая раковина — это самостоятельная кость черепа.
Шовный материал — неподвижный сустав между соседними костями черепа. Узкий промежуток между костями заполнен плотной волокнистой соединительной тканью, соединяющей кости.Длинные швы, расположенные между костями черепа, не прямые, а вместо этого идут по неправильным, сильно изгибающимся путям. Эти изгибающиеся линии служат для того, чтобы плотно сцепить соседние кости, тем самым добавляя прочности черепу для защиты мозга.
Две линии швов, видимые на верхней части черепа, представляют собой коронарный и сагиттальный швы. Венечный шов проходит из стороны в сторону поперек черепа в коронарной плоскости сечения (см. Рисунок 7.3.3). Он соединяет лобную кость с правой и левой теменными костями.Сагиттальный шов проходит кзади от коронарного шва в месте пересечения, называемого bregma , и проходит по средней линии в верхней части черепа в сагиттальной плоскости разреза (см. Рисунок 7.3.8). Он объединяет правую и левую теменные кости. На задней части черепа сагиттальный шов заканчивается присоединением лямбдовидного шва в месте пересечения, которое называется лямбда . Лямбдовидный шов проходит вниз и латерально в обе стороны от места соединения со сагиттальным швом.Лямбдовидный шов соединяет затылочную кость с правой и левой теменными и височными костями. Этот шов назван в честь его перевернутой буквы «V», которая напоминает заглавную версию греческой лямбды (Λ). Плоский шов расположен на боковой поверхности черепа. Он соединяет плоскоклеточную часть височной кости с теменной костью (см. Рисунок 7.3.3). На пересечении лобной кости, теменной кости, плоской части височной кости и большого крыла клиновидной кости находится птерион , небольшая линия шва в форме буквы «Н», соединяющая область.Это самая слабая часть черепа. Птерион расположен примерно на два пальца выше скуловой дуги и на ширине большого пальца кзади от верхней части скуловой кости.
Рисунок 7.3.12 Швы черепа
Заболевания… скелетной системы
Головные и черепно-мозговые травмы являются основными причинами немедленной смерти и инвалидности, а кровотечение и инфекции являются возможными дополнительными осложнениями. По данным Центров по контролю и профилактике заболеваний (2010 г.), примерно 30 процентов всех смертей, связанных с травмами, в США вызваны травмами головы.Большинство травм головы связаны с падениями. Они наиболее распространены среди детей младшего возраста (возраст 0–4 года), подростков (15–19 лет) и пожилых людей (старше 65 лет). Дополнительные причины различаются, но наиболее заметными из них являются автомобильные и мотоциклетные аварии.
Сильные удары по черепу могут вызвать переломы. Это может привести к кровотечению внутри черепа с последующим повреждением головного мозга. Наиболее распространенным является линейный перелом черепа, при котором линии перелома расходятся от точки удара.Другие типы переломов включают оскольчатый перелом, при котором кость разбивается на несколько частей в месте удара, или вдавленный перелом, при котором сломанная кость проталкивается внутрь. При переломе контратакой (встречным ударом) кость в месте удара не ломается, а вместо этого происходит перелом на противоположной стороне черепа. Таким образом, могут возникнуть переломы затылочной кости у основания черепа, в результате чего возникает базилярный перелом, который может повредить артерию, проходящую через сонный канал.
Удар по боковой стороне головы может привести к перелому костей птериона. Птерион является важным клиническим ориентиром, потому что расположенный непосредственно глубоко внутри черепа — это основная ветвь артерии, которая снабжает череп и покрывающие слои мозга. Сильный удар в эту область может сломать кости вокруг птериона. Если основная артерия повреждена, кровотечение может вызвать образование гематомы (скопление крови) между мозгом и внутренней частью черепа.По мере того как кровь накапливается, она оказывает давление на мозг. Симптомы, связанные с гематомой, могут не проявиться сразу после травмы, но при отсутствии лечения скопление крови будет оказывать повышенное давление на мозг и может привести к смерти в течение нескольких часов.
Внешний веб-сайт
Просмотрите эту анимацию, чтобы увидеть, как удар по голове может привести к обратному перелому (ответный удар) базилярной части затылочной кости на основании черепа.Почему базилярный перелом может быть опасным для жизни?
Лицевые кости черепа образуют верхнюю и нижнюю челюсти, нос, носовую полость и носовую перегородку, а также глазницу. Лицевые кости включают 14 костей, из которых шесть парных и две непарных. Парные кости — это верхняя, небная, скуловая, носовая, слезная и нижняя носовые раковины. Непарные кости — это кости сошника и нижней челюсти. Хотя решетчатая кость классифицируется как черепные кости, она также способствует развитию носовой перегородки и стенок носовой полости и глазницы.
Верхнечелюстная кость
Верхнечелюстная кость , часто называемая просто верхней челюстью (множественное число = maxillae), является одной из пары, которые вместе образуют верхнюю челюсть, большую часть твердого неба, медиальное дно орбиты и латеральное основание нос (см. рисунок 7.3.2). Изогнутый нижний край верхнечелюстной кости, который образует верхнюю челюсть и содержит верхние зубы, представляет собой альвеолярный отросток верхней челюсти (рис. 7.3.13). Каждый зуб закреплен в глубокой лунке, называемой альвеолой.На передней верхней челюсти, чуть ниже глазницы, находится подглазничное отверстие. Это точка выхода сенсорного нерва, который снабжает нос, верхнюю губу и переднюю щеку. На нижнем черепе можно увидеть небный отросток от каждой верхнечелюстной кости, соединяющийся вместе по средней линии, образуя передние три четверти твердого неба (см. Рис. 7.3.7 a ). Твердое небо — это костная пластинка, которая образует верхнюю часть рта и дно носовой полости, разделяя ротовую и носовую полости.
Рисунок 7.3.13 — Верхнечелюстная кость: Верхнечелюстная кость образует верхнюю челюсть и поддерживает верхние зубы. Каждая верхняя челюсть также образует латеральное дно каждой глазницы и большую часть твердого неба.
Небная кость
Небная кость — одна из пары костей неправильной формы, которые вносят небольшие участки в боковые стенки носовой полости и медиальную стенку каждой глазницы. Самая большая область каждой небной кости — это горизонтальная пластина .Пластины правой и левой небных костей соединяются по средней линии, образуя заднюю четверть твердого неба (см. Рис. 7.3.7 a ). Таким образом, небные кости лучше всего видны на нижнем виде черепа и твердого неба.
Гомеостатический дисбаланс… Расщелина губы и неба
Во время эмбрионального развития правая и левая верхнечелюстные кости соединяются по средней линии, образуя верхнюю челюсть. В то же время мышцы и кожа, покрывающие эти кости, соединяются вместе, образуя верхнюю губу.Внутри рта небные отростки костей верхней челюсти вместе с горизонтальными пластинами правой и левой небных костей соединяются вместе, образуя твердое небо. Если в этих процессах развития возникает ошибка, это может привести к врожденному дефекту расщелины губы или неба.
Заячья губа — это распространенный порок развития, который встречается примерно у 1: 1000 новорожденных, большинство из которых — мужчины. Этот дефект заключается в частичном или полном отказе правой и левой частей верхней губы срастаться друг с другом, в результате чего остается расщелина (щель).
Более серьезный порок развития — волчья пасть, поражающая твердое небо. Твердое небо — это костная структура, отделяющая носовую полость от ротовой полости. Он образуется во время эмбрионального развития путем слияния небных и верхнечелюстных костей по средней линии. Расщелина неба встречается примерно у 1: 2500 рождений и чаще встречается у женщин. Это результат неспособности двух половин твердого неба полностью соединиться и слиться по средней линии, в результате чего между ними остается промежуток.Этот промежуток обеспечивает сообщение между носовой и ротовой полостями. В тяжелых случаях костная щель продолжается до передней верхней челюсти, где альвеолярные отростки костей верхней челюсти также не соединяются должным образом над передними зубами. Если это произойдет, также будет видна заячья губа. Из-за сообщения между ротовой и носовой полостями, расщелина неба очень затрудняет для младенца процесс сосания, необходимый для кормления, что подвергает его риску недоедания.Для исправления дефекта волчьей пасти требуется хирургическое вмешательство.
Скуловая кость
скуловая кость также известна как скула. Каждая из парных скуловых костей образует большую часть боковой стенки глазницы и боковых-нижних краев переднего отверстия глазницы (см. Рисунок 7.3.2). Короткий височный отросток скуловой кости выступает кзади, где он образует переднюю часть скуловой дуги (см. Рисунок 7.3.3).
Носовая кость
Носовая кость — одна из двух маленьких костей, которые соединяются друг с другом, образуя костную основу (переносицу) носа.Они также поддерживают хрящи, образующие боковые стенки носа (см. Рисунок 7.3.10). Это кости, которые повреждаются при переломе носа.
Слезная кость
Каждая слезная кость представляет собой небольшую прямоугольную кость, которая формирует переднемедиальную стенку глазницы (см. Рисунок 7.3.2 и Рисунок 7.3.3). Передняя часть слезной кости образует неглубокое углубление, называемое слезной ямкой , а снизу от него идет носослезный канал .Слезная жидкость (слезы глаза), которая поддерживает влажную поверхность глаза, стекает через медиальный угол глаза в носослезный канал. Затем этот проток расширяется вниз и открывается в носовую полость за нижними носовыми раковинами. В носовой полости слезная жидкость обычно оттекает кзади, но при усилении слезотечения из-за плача или раздражения глаз некоторое количество жидкости также будет стекать кпереди, вызывая насморк.
Нижние носовые раковины
Правая и левая нижние носовые раковины образуют изогнутую костную пластинку (носовую раковину), которая выступает в пространство носовой полости от нижней боковой стенки (см.рисунок 7.3.12). Нижняя раковина является самой большой из носовых раковин, и ее легко увидеть, если заглянуть в переднее отверстие носовой полости.
Вомер кость
Непарный сошник, часто называемый просто сошником, имеет треугольную форму и образует задне-нижнюю часть носовой перегородки (см. Рисунок 7.3.10). Сошник лучше всего видно, если смотреть сзади в задние отверстия носовой полости (см. Рис. 7.3.7 a ). На этом изображении видно, что сошник формирует носовую перегородку по всей высоте.Если смотреть в переднее отверстие носовой полости, можно также увидеть гораздо меньшую часть сошника.
Нижняя челюсть
Нижняя челюсть образует нижнюю челюсть и является единственной подвижной костью черепа. Во время рождения нижняя челюсть состоит из парных правой и левой костей, но в течение первого года они сливаются вместе, образуя единую U-образную нижнюю челюсть взрослого черепа. Каждая сторона нижней челюсти состоит из горизонтального тела и сзади вертикально ориентированной ветви нижней челюсти (ветвь = «ветвь»).Внешний край нижней челюсти, где соединяются тело и ветвь, называется углом нижней челюсти (рис. 7.3.14).
Ветви на каждой стороне нижней челюсти имеют два восходящих костных выступа. Более передний выступ — это уплощенный венечный отросток нижней челюсти , который обеспечивает прикрепление одной из кусающих мышц. Задний выступ представляет собой мыщелок нижней челюсти , который завершается мыщелком овальной формы .Мыщелок нижней челюсти сочленяется (соединяется) с нижнечелюстной ямкой и суставным бугорком височной кости. Вместе эти сочленения образуют височно-нижнечелюстной сустав, который позволяет открывать и закрывать рот (см. Рисунок 7.3.3). Широкая U-образная кривая, расположенная между венечным и мыщелковым отростками, представляет собой вырез нижней челюсти .
Важные ориентиры для нижней челюсти включают следующее:
- Альвеолярный отросток нижней челюсти — Это верхняя граница тела нижней челюсти, служащая для фиксации нижних зубов.
- Ментальный выступ —Передняя проекция нижнего края передней нижней челюсти, которая образует подбородок (подбородок = подбородок).
- Ментальное отверстие —Отверстие, расположенное с каждой стороны переднебоковой нижней челюсти, которое является местом выхода сенсорного нерва, снабжающего подбородок.
- Мило-подъязычная линия — Этот костный гребень проходит вдоль внутренней части тела нижней челюсти (см. Рисунок 7.3.10). Мышца, образующая дно ротовой полости, прикрепляется к подъязычным линиям с обеих сторон нижней челюсти.
- Отверстие нижней челюсти — Это отверстие расположено на медиальной стороне ветви нижней челюсти. Отверстие ведет в туннель, который проходит по всей длине тела нижней челюсти. Чувствительный нерв и кровеносные сосуды, снабжающие нижние зубы, входят в отверстие нижней челюсти и затем следуют по этому туннелю. Таким образом, чтобы обезболить нижние зубы перед стоматологической работой, стоматолог должен ввести анестезию в боковую стенку ротовой полости в точке, предшествующей тому, где этот чувствительный нерв входит в отверстие нижней челюсти.
- Lingula — Этот небольшой костный лоскут назван в честь своей формы (lingula = «маленький язычок»). Он расположен непосредственно рядом с отверстием нижней челюсти, на медиальной стороне ветви. Связка, фиксирующая нижнюю челюсть во время открывания и закрывания рта, идет вниз от основания черепа и прикрепляется к язычку.
Рисунок 7.3.14 — Изолированная нижняя челюсть: Нижняя челюсть — единственная подвижная кость черепа.
Орбита — это костная впадина, в которой находится глазное яблоко и мышцы, которые двигают глазное яблоко или открывают верхнее веко.Каждая глазница имеет конусовидную форму с узкой задней частью, которая расширяется к большому переднему отверстию. Чтобы защитить глаз, костные края переднего отверстия утолщены и несколько сужены. Медиальные стенки двух орбит параллельны друг другу, но каждая боковая стенка отклоняется от средней линии под углом 45 °. Это расхождение обеспечивает большее боковое периферическое зрение.
Стенки каждой орбиты включают в себя семь костей черепа (рис. 7.3.15).Лобная кость образует крышу, а скуловая кость образует боковую стенку и боковое дно. Медиальное дно в основном образовано верхней челюстью с небольшой долей небной кости. Решетчатая кость и слезная кость составляют большую часть медиальной стенки, а клиновидная кость образует заднюю орбиту.
На задней вершине орбиты находится отверстие зрительного канала , через которое зрительный нерв проходит от сетчатки к головному мозгу. Сбоку от него находится удлиненная верхняя глазничная щель неправильной формы, которая обеспечивает проход для артерии, снабжающей глазное яблоко, сенсорных нервов и нервов, снабжающих мышцы, участвующие в движениях глаз.
Рисунок 7.3.15 — Кости орбиты: Семь костей черепа составляют стенки орбиты. Из полости черепа в заднюю орбиту выходят зрительный канал и верхняя глазничная щель.
Носовая перегородка состоит как из кости, так и из хряща (рисунок 7.3.16; см. Также рисунок 7.3.10). Верхняя часть перегородки образована перпендикулярной пластиной решетчатой кости. Нижняя и задняя части перегородки образованы сошниковой костью треугольной формы.Передняя перегородка носа образована перегородочным хрящом , гибкой пластиной, заполняющей промежуток между перпендикулярной пластинкой решетчатой и сошниковой костей. Этот хрящ также выходит наружу в нос, где разделяет правую и левую ноздри.
К боковой стенке с каждой стороны носовой полости прикреплены верхняя, средняя и нижняя носовые раковины (singular = concha), названные по их положению (см. Рисунок 7.3.12). Это костные пластинки, которые изгибаются вниз, выступая в носовую полость.Они служат для закрутки входящего воздуха, что помогает согреть и увлажнить его, прежде чем воздух попадет в нежные воздушные мешочки легких. Это также позволяет слизи, выделяемой тканью, выстилающей носовую полость, улавливать поступающую пыль, пыльцу, бактерии и вирусы. Самыми крупными из раковин являются нижние носовые раковины, которые представляют собой независимую кость черепа. Средние и самые маленькие раковины образованы решетчатой костью. При взгляде в переднее носовое отверстие черепа можно увидеть только нижнюю и среднюю раковины.Небольшие верхние носовые раковины хорошо скрыты над средними раковинами и за ними.
Рисунок 7.3.16 — Носовая перегородка: Носовая перегородка образована перпендикулярной пластиной решетчатой кости и сошниковой костью. Перегородочный хрящ заполняет промежуток между этими костями и простирается в нос.
придаточные пазухи носа представляют собой полые, заполненные воздухом пространства, расположенные внутри определенных костей черепа (рис. 7.3.17). Все пазухи сообщаются с полостью носа (околоносовые = «рядом с полостью носа») и выстланы слизистой оболочкой носа.Они служат для уменьшения костной массы и, таким образом, осветления черепа, а также добавляют резонанса голосу. Эта вторая особенность наиболее очевидна, когда у вас простуда или заложенность носовых пазух, которые вызывают отек слизистой оболочки и избыточное выделение слизи, закупоривая узкие проходы между пазухами и носовой полостью и заставляя ваш голос звучать иначе, чем вы и другие. Эта закупорка также может позволить пазухам заполниться жидкостью, в результате чего давление будет вызывать боль и дискомфорт.
Придаточные пазухи носа названы в честь кости черепа, которую каждая занимает. Лобная пазуха расположена чуть выше бровей, внутри лобной кости (см. Рисунок 7.3.16). Это нерегулярное пространство может быть разделено по средней линии на двусторонние пространства или они могут быть объединены в одно пространство синуса. Лобная пазуха — это самый передний из придаточных пазух носа. Самая большая пазуха — гайморовая пазуха . Они парные и расположены в правой и левой верхнечелюстных костях, где они занимают область чуть ниже глазниц.При инфекциях носовых пазух чаще всего поражаются верхнечелюстные пазухи. Поскольку их соединение с носовой полостью расположено высоко на их медиальной стенке, их трудно дренировать. Клиновидная пазуха представляет собой единую срединную пазуху. Он расположен в теле клиновидной кости, непосредственно впереди и ниже турецкого седла, что делает его самым задним из придаточных пазух носа. На боковых сторонах решетчатой кости есть несколько небольших пространств, разделенных очень тонкими костными стенками.Каждое из этих пространств называется решетчатой воздушной ячейкой . Они расположены по обе стороны от решетчатой кости, между верхней носовой полостью и медиальной орбитой, сразу за верхними носовыми раковинами.
Рисунок 7.3.17 — Параназальные пазухи: Заполненные воздухом придаточные пазухи носа, каждая из которых названа в честь кости, в которой они находятся, стекают в носовую полость.
Подъязычная кость — это независимая кость, которая не контактирует с другими костями и, следовательно, не является частью черепа (рис. 7.3.18). Это небольшая U-образная кость, расположенная в верхней части шеи на уровне нижней челюсти, с кончиками буквы U, направленной назад. Подъязычная кость служит основанием для языка вверху и прикрепляется к гортани внизу и к глотке сзади. Подъязычная кость удерживается на месте серией маленьких мышц, которые прикрепляются к ней сверху или снизу. Эти мышцы действуют для перемещения подъязычной кости вверх / вниз или вперед / назад. Движения подъязычной кости координируются с движениями языка, гортани и глотки при глотании и разговоре.
Рисунок 7.3.18 — Подъязычная кость: Подъязычная кость расположена в верхней части шеи и не соединяется с другими костями. Он обеспечивает прикрепление мышц к языку, гортани и глотке.
Обзор главы
Череп состоит из черепа и лицевых костей. Череп окружает и защищает мозг, который занимает полость черепа внутри черепа. Он состоит из округлого свода черепа и сложного основания. Череп состоит из восьми костей: парных теменных и височных костей, а также непарных лобных, затылочных, клиновидных и решетчатых костей.Узкий промежуток между костями заполнен плотной волокнистой соединительной тканью, соединяющей кости. Сагиттальный шов соединяет правую и левую теменные кости. Венечный шов соединяет теменные кости с лобной костью, лямбдовидный шов соединяет их с затылочной костью, а плоский шов соединяет их с височной костью. Дно полости черепа увеличивается в глубину спереди назад и делится на три черепные ямки; передняя черепная ямка, средняя черепная ямка и задняя черепная ямка.
Кости лица поддерживают структуры лица и образуют верхнюю и нижнюю челюсти. Они состоят из 14 костей, включая парные верхнечелюстные, небные, скуловые, носовые, слезные и нижние раковины, а также непарные сошковые и нижние челюсти. Решетчатая кость также способствует формированию структур лица. Верхняя челюсть образует верхнюю челюсть, а нижняя челюсть — нижнюю. Верхняя челюсть также образует большую переднюю часть твердого неба, которая завершается меньшими небными костями, которые образуют заднюю часть твердого неба.
Передняя часть черепа имеет орбиты, в которых расположены глазные яблоки и связанные с ними мышцы. Стенки орбиты образованы семью костями: лобной, скуловой, верхней, небной, решетчатой, слезной и клиновидной. На верхнем крае орбиты находится надглазничное отверстие, а под глазницей — подглазничное отверстие. Нижняя челюсть имеет два отверстия: нижнечелюстное отверстие на внутренней поверхности и подбородочное отверстие на внешней поверхности возле подбородка.Раковины носа — это костные выступы боковых стенок полости носа. Большая нижняя носовая раковина является самостоятельной костью, а средняя и верхняя раковины являются частями решетчатой кости. Носовая перегородка образована перпендикулярной пластиной решетчатой кости, сошниковой костью и перегородочным хрящом. Придаточные пазухи представляют собой заполненные воздухом пространства, расположенные внутри лобной, верхнечелюстной, клиновидной и решетчатой костей.
На боковой поверхности черепа скуловая дуга состоит из двух частей: височного отростка скуловой кости спереди и скулового отростка височной кости сзади.Височная ямка — это неглубокое пространство, расположенное на боковой поверхности черепа над уровнем скуловой дуги. Подвисочная ямка расположена ниже скуловой дуги и глубоко до ветви нижней челюсти.
Подъязычная кость расположена в верхней части шеи и не соединяется ни с одной другой костью. Он удерживается мышцами и служит опорой для языка вверху, гортани внизу и глотки сзади.
Вопросы по интерактивной ссылке
Посмотрите это видео, чтобы увидеть вращающийся взорванный череп с разноцветными костями.Какая кость (желтая) расположена в центре и соединяется с большинством других костей черепа?
Клиновидная кость соединяется с большинством других костей черепа. Он расположен в центре, где он образует части округлой оболочки мозга и основания черепа.
Просмотрите эту анимацию, чтобы увидеть, как удар по голове может привести к обратному перелому (ответный удар) базилярной части затылочной кости на основании черепа. Почему базилярный перелом может быть опасным для жизни?
Перелом базиляра может повредить артерию, входящую в череп, вызывая кровотечение в головном мозге.
Обзорные вопросы
Вопросы о критическом мышлении
1. Определите и перечислите кости, которые образуют череп или поддерживают лицевые структуры.
2. Определите основные швы черепа, их расположение и кости, соединенные каждым из них.
3. Опишите переднюю, среднюю и заднюю черепные ямки.
4. Опишите части носовой перегородки как в сухом, так и в живом черепе.
Центры по контролю и профилактике заболеваний (США). Профилактика травм и борьба с ними: черепно-мозговые травмы [Интернет]. Атланта, Джорджия; [цитируется 18 марта 2013 г.].
Доступно по адресу: http://www.cdc.gov/traumaticbraininjury/statistics.html.
Глоссарий
- альвеолярный отросток нижней челюсти
- верхняя граница тела нижней челюсти, содержащая нижние зубы
- альвеолярный отросток верхней челюсти
- изогнутый нижний край верхней челюсти, который поддерживает и фиксирует верхние зубы
- угол нижней челюсти
- закругленный угол, расположенный на внешнем краю соединения тела и ветви
- передняя черепная ямка
- Самая мелкая и наиболее передняя черепная ямка основания черепа, которая простирается от лобной кости до малого крыла клиновидной кости
- суставной бугорок
- гладкий гребень, расположенный на нижней части черепа, непосредственно перед нижней челюстной ямкой
- кальвария
- (также тюбетейка) закругленная вершина черепа
- сонный канал
- зигзагообразный туннель, обеспечивающий проход внутренней сонной артерии через основание черепа к головному мозгу; начинается перед шиловидным отростком и заканчивается в средней полости черепа, около заднебокового основания турецкого седла
- мыщелковый отросток нижней челюсти
- утолщение кверху от заднего края ветви нижней челюсти
- мыщелок
- овальный отросток, расположенный вверху мыщелкового отростка нижней челюсти
- коронковый шов
- сустав, соединяющий лобную кость с правой и левой теменными костями через верхнюю часть черепа
- венечный отросток нижней челюсти
- Уплощенный выступ вверх от переднего края ветви нижней челюсти
- полость черепа
- внутреннее пространство черепа, в котором находится мозг
- череп
- Часть черепа, окружающая мозг
- решетчатая пластина
- небольшие уплощенные участки с множеством мелких отверстий, расположенные по обе стороны от средней линии в дне передней черепной ямки; образована решетчатой костью
- Криста Галли
- небольшой выступ вверх, расположенный по средней линии в дне передней черепной ямки; образована решетчатой костью
- решетчатая воздушная ячейка
- одно из нескольких небольших заполненных воздухом пространств, расположенных на боковых сторонах решетчатой кости, между глазницей и верхней полостью носа
- решетчатая кость
- непарная кость, которая образует крышу и верхнюю боковую стенку полости носа, части дна передней черепной ямки и медиальную стенку орбиты, а также верхнюю часть носовой перегородки
- наружный слуховой проход
- отверстие слухового прохода, расположенное на боковой стороне черепа
- наружный затылочный бугор
- небольшая бугорка, расположенная по средней линии задней части черепа
- лицевые кости
- четырнадцать костей, которые поддерживают структуры лица и образуют верхнюю и нижнюю челюсти и твердое небо
- кружевное отверстие
- отверстие неправильной формы в основании черепа, расположенное ниже выхода сонного канала
- большое затылочное отверстие
- большое отверстие в затылочной кости черепа, через которое выходит спинной мозг и позвоночные артерии входят в череп
- овальное отверстие средней черепной ямки
- отверстие овальной формы в дне средней черепной ямки
- круглое отверстие
- круглое отверстие в дне средней черепной ямки, расположенное между верхней глазничной щелью и овальным отверстием
- остистое отверстие
- небольшое отверстие в дне средней черепной ямки, расположенное латеральнее овального отверстия
- лобная кость
- Непарная кость, образующая лоб, крышу орбиты и дно передней черепной ямки
- лобная пазуха
- пространство лобной кости заполненное воздухом; передняя часть придаточных пазух носа
- глабель
- небольшое вдавление лобной кости, расположенное по средней линии между бровями
- большие крылья клиновидной кости
- боковые проекции клиновидной кости, образующие переднюю стенку средней черепной ямки и область боковой черепа
- твердое небо
- Костная структура, образующая верхнюю часть рта и дно носовой полости, образованная небным отростком верхнечелюстных костей и горизонтальной пластиной небных костей
- пластина горизонтальная
- медиальное продолжение небной кости, которое образует заднюю четверть твердого неба
- подъязычный канал
- парные отверстия, которые проходят кпереди от переднебокового края большого затылочного отверстия глубоко к затылочным мыщелкам
- гипофизарная (гипофизарная) ямка
- неглубокое углубление на вершине турецкого седла, в котором находится гипофиз (гипофиз)
- нижняя носовая раковина
- одна из парных костей, которые выступают из боковых стенок носовой полости и образуют самую большую и самую нижнюю из носовых раковин
- подглазничное отверстие
- отверстие на передней части черепа, ниже глазницы
- подвисочная ямка
- пространство на боковой стороне черепа, ниже уровня скуловой дуги и глубоко (медиально) до ветви нижней челюсти
- внутренний слуховой проход
- отверстие в каменистом гребне, расположенном на боковой стенке задней черепной ямки
- яремное отверстие
- отверстие неправильной формы, расположенное в боковом дне задней полости черепа
- слезная кость
- парных костей, образующих передне-медиальную стенку каждой глазницы
- слезная ямка
- неглубокое углубление в передне-медиальной стенке глазницы, образованное слезной костью, дающей начало носослезному каналу
- лямбдовидный шов
- перевернутый V-образный сустав, соединяющий затылочную кость с правой и левой теменными костями на задней части черепа
- пластина латеральная крыловидная
- парные уплощенные костные выступы клиновидной кости, расположенные на нижнем черепе, латеральнее медиальной крыловидной пластинки
- малые крылья клиновидной кости
- боковые отростки клиновидной кости, образующие костную губу, разделяющую переднюю и среднюю черепные ямки
- язык
- небольшой костный лоскут, расположенный на внутренней (медиальной) поверхности ветви нижней челюсти, рядом с отверстием нижней челюсти
- нижняя челюсть
- кость непарная, образующая кость нижней челюсти; единственная подвижная кость черепа
- нижнечелюстное отверстие
- отверстие, расположенное на внутренней (медиальной) поверхности ветви нижней челюсти
- нижнечелюстная ямка
- овальное углубление на нижней поверхности черепа
- вырез на нижней челюсти
- большая U-образная выемка, расположенная между мыщелковым отростком и венечным отростком нижней челюсти
- сосцевидный отросток
- большой костный выступ на нижней боковой поверхности черепа сразу за мочкой уха
- верхнечелюстная кость
- (также верхняя челюсть) парные кости, образующие верхнюю челюсть и переднюю часть твердого неба
- гайморовая пазуха
- воздушное пространство, расположенное с каждой верхнечелюстной костью; самая большая из придаточных пазух носа
- пластина крыловидная медиальная
- парные уплощенные костные выступы клиновидной кости, расположенные на нижней части черепа медиальнее латеральной крыловидной пластинки; образуют заднюю часть боковой стенки полости носа
- подбородочное отверстие
- отверстие, расположенное на переднебоковой стороне тела нижней челюсти
- подбородочная выпуклость
- нижний край передней нижней челюсти, образующий подбородок
- средняя черепная ямка
- Центрально расположенная черепная ямка, которая простирается от малых крыльев клиновидной кости до каменистого гребня
- средняя носовая раковина
- носовая раковина, образованная решетчатой костью, расположенной между верхней и нижней раковинами
- миелоидная линия
- Костный гребень, расположенный по внутренней (медиальной) поверхности тела нижней челюсти
- носовая кость
- парные кости, образующие основу носа
- полость носа
- отверстие в черепе для прохода воздуха
- носовые раковины
- костные пластинки изогнутые, выступающие из боковых стенок носовой полости; включают верхнюю и среднюю носовые раковины, которые являются частями решетчатой кости, и независимую нижнюю носовую раковину
- носовая перегородка
- плоская срединная структура, разделяющая носовую полость на половины, образованная перпендикулярной пластиной решетчатой кости, сошниковой костью и перегородочным хрящом
- носослезный канал
- проход для дренирования слез, который идет вниз от медиально-передней орбиты в носовую полость, заканчиваясь позади нижних носовых раковин
- затылочная кость
- непарная кость, которая образует задние части оболочки мозга и основание черепа
- затылочный мыщелок
- парные костные выступы овальной формы, расположенные на нижней части черепа по обе стороны от большого затылочного отверстия
- оптический канал
- отверстие между средней черепной ямкой и задней орбитой
- орбита
- костная впадина, которая содержит глазное яблоко и связанные с ним мышцы
- небная кость
- парные кости, образующие заднюю четверть твердого неба и небольшой участок дна глазницы
- небный отросток
- медиальный выступ верхней челюсти, образующий передние три четверти твердого неба
- придаточные пазухи носа
- полости черепа, соединенные с раковинами, которые служат для обогрева и увлажнения поступающего воздуха, выделения слизи и уменьшения веса черепа; состоят из лобных, верхнечелюстных, клиновидных и решетчатых пазух
- теменная кость
- парные кости, образующие верхнюю, боковую стороны черепа
- пластинка перпендикулярная решетчатой кости
- вниз, продолжение по средней линии решетчатой кости, которая образует верхнюю часть носовой перегородки
- каменистый гребень
- каменистая часть височной кости, которая образует большой треугольный гребень в дне полости черепа, разделяющий среднюю и заднюю черепные ямки; вмещает структуры среднего и внутреннего уха
- задняя черепная ямка
- самая глубокая и самая задняя черепная ямка; простирается от каменистого гребня до затылочной кости
- птерион
- Н-образная область соединения швов, соединяющая лобную, теменную, височную и клиновидную кости на боковой стороне черепа
- ветвь нижней челюсти
- вертикальная часть нижней челюсти
- сагиттальный шов
- сустав, соединяющий правую и левую теменные кости по средней линии вдоль верхней части черепа
- Селла турчика
- приподнятая область клиновидной кости, расположенная по средней линии средней черепной ямки
- перегородочный хрящ
- Плоская хрящевая структура, образующая переднюю часть носовой перегородки
- череп
- вместе черепные и челюстно-лицевые кости
- клиновидная кость
- непарная кость, образующая центральное основание черепа
- клиновидная пазуха
- воздушное пространство, расположенное внутри клиновидной кости; самый задний из придаточных пазух носа
- плоский шов
- сустав, соединяющий теменную кость с плоской частью височной кости на боковой стороне черепа
- шиловидный отросток
- выступающий вниз удлиненный костный отросток, расположенный на нижней поверхности черепа
- шилососцевидное отверстие
- отверстие, расположенное на нижней части черепа, между шиловидным отростком и сосцевидным отростком
- верхняя носовая раковина
- наименьшая и наиболее высоко расположенная из носовых раковин; образована решетчатой костью
- верхняя затылочная линия
- парные костные линии на задней части черепа, которые отходят латерально от наружного затылочного выступа
- верхняя глазничная щель
- отверстие неправильной формы между средней черепной ямкой и задней орбитой
- надглазничное отверстие
- отверстие на передней части черепа, у верхнего края глазницы
- надглазничный край
- верхний край орбиты
- шовный
- линия соединения, на которой соседние кости черепа объединены фиброзной соединительной тканью
- височная кость
- парные кости, образующие боковые, нижние части черепа, с чешуйчатой, сосцевидной и каменистой частями
- височная ямка
- мелкое пространство на боковой стороне черепа выше уровня скуловой дуги
- височный отросток скуловой кости
- короткий отросток от скуловой кости, образующий переднюю часть скуловой дуги
- сошковая кость
- непарная кость, образующая нижнюю и заднюю части носовой перегородки
- скуловая дуга
- Удлиненная свободно стоящая дуга на боковой поверхности черепа, образованная спереди височным отростком скуловой кости, а сзади скуловым отростком височной кости
- скуловая кость
- скула; парные кости, образующие боковую орбиту и переднюю скуловую дугу
- скуловой отросток височной кости
- расширение от височной кости, которое образует заднюю часть скуловой дуги
Решения
Ответы на вопросы о критическом мышлении
- Мозговой футляр — это часть черепа, которая окружает и защищает мозг.Он подразделяется на округлую вершину черепа, называемую сводом черепа, и основание черепа. Корпус мозга состоит из восьми костей. Это парные теменные и височные кости, а также непарные лобная, затылочная, клиновидная и решетчатая кости. Кости лица поддерживают структуры лица и образуют верхнюю и нижнюю челюсти, носовую полость, носовую перегородку и глазницу. Всего 14 лицевых костей. Это парные верхнечелюстные, небные, скуловые, носовые, слезные и нижние носовые раковины, а также непарные сошковые и нижнечелюстные кости.
- Венечный шов проходит через верхнюю часть передней части черепа. Он соединяет переднюю лобную кость с правой и левой теменными костями. Сагиттальный шов проходит по средней линии на верхней части черепа. Он соединяет между собой правую и левую теменные кости. Плоский шов представляет собой изогнутый шов, расположенный на боковой стороне черепа. Он соединяет плоскоклеточную часть височной кости с теменной костью. Лямбдовидный шов расположен на задней части черепа и имеет перевернутую V-образную форму.Он соединяет затылочную кость с правой и левой теменными костями.
- Передняя черепная ямка — самая мелкая из трех черепных ямок. Он простирается от лобной кости кпереди до малого крыла клиновидной кости кзади. Он разделен по средней линии crista galli и решетчатыми пластинами решетчатой кости. Средняя черепная ямка расположена в центральном черепе и глубже передней ямки. Средняя ямка проходит от малого крыла клиновидной кости спереди до каменистого гребня сзади.По средней линии он разделен турецким седлом. Задняя черепная ямка — самая глубокая ямка. Он простирается от каменистого гребня спереди до затылочной кости сзади. Большое затылочное отверстие расположено по средней линии задней ямки.
- В сухом черепе две костные части носовой перегородки. Перпендикулярная пластинка решетчатой кости образует верхнюю часть перегородки. Сошниковая кость образует нижнюю и заднюю части перегородки. В живом черепе перегородочный хрящ завершает перегородку, заполняя переднюю часть между костными компонентами и распространяясь наружу в нос.
Анатомия гортани: хрящи, связки и мышцы
Автор:
Эдвин Окран МБЧБ, магистр наук
•
Рецензент:
Димитриос Митилинайос MD, PhD
Последний раз отзыв: 29 октября 2020 г.
Время чтения: 13 минут
Гортань (вид спереди)
Гортань представляет собой сложную полую структуру, расположенную в передней средней линии шеи. Он находится кпереди от пищевода и на уровне третьего-шестого шейных позвонков в нормальном положении.Он состоит из хрящевого скелета, соединенного мембранами, связками и связанными с ними мышцами, которые удерживают его от окружающих структур. Он расположен чуть выше трахеи и продолжается выше ротоглотки (часть глотки позади ротовой полости).
Гортань направляет воздух в нижние дыхательные пути и закрывает дыхательные пути, особенно во время глотания, чтобы предотвратить всасывание пищи. Его обычно называют «голосовой ящик» или «орган звучания» , так как в нем находится структура, отвечающая за производство звука.Он довольно подвижен в шее, и его можно увидеть и почувствовать, когда он движется вверх и вперед во время глотания, закрывая трахею и открывая пищевод.
| Функции | Воздуховод, защита дыхательных путей, производство звука |
| Хрящи | Непарные (3): перстневидный рубец, щитовидная железа и надгортанник Парные (3): черноволосый, роговой и клинописный |
| Полость | Три части: преддверие, средняя часть, подглоточная полость Glottis: голосовые связки, rima glottidis |
| Мышцы | Внешний: движение всей гортани Внутренний: движение компонентов гортани |
| Кровоснабжение | Верхняя и нижняя гортанные артерии, верхняя и нижняя гортанные вены |
| Лимфодренаж | Верхние и нижние глубокие шейные лимфатические узлы |
| Иннервация | Блуждающий нерв: верхние гортанные нервы (внутренний и внешний), нижние гортанные нервы |
В этой статье будут рассмотрены основные анатомические структуры и основные функции гортани.Он также описывает кровоснабжение, иннервацию и лимфодренаж гортани, а также клиническое применение и интересный факт.
Хрящевой каркас и связки
Гортань состоит из трех крупных непарных хрящей (перстневидный, щитовидный и надгортанник) и трех парных более мелких хрящей (черпаловидный, роговой и клиновидный), составляющих в общей сложности девять отдельных хрящей. щитовидный хрящ является самым большим из хрящей гортани и состоит из гиалинового хряща.Он образует переднюю и боковую части гортани и не имеет заднего компонента. Широкие плоские правая и левая половины (laminae) хряща сливаются спереди по средней линии, образуя V-образный передний выступ, называемый гортанным выступом (обычно называемым «кадык»).
Адамово яблоко обычно более заметно у мужчин после полового созревания. Это связано с влиянием гормона тестостерона, который стимулирует общий рост гортани, что со временем приводит к более глубокому голосу.Выше этого выступа находится верхняя щитовидная железа выемка . Менее заметная нижняя вырезка щитовидной железы присутствует вдоль основания щитовидного хряща. Задний край каждой пластинки простирается вверх в верхний рог и вниз в нижний рог . Более длинный верхний рог вместе со всей верхней границей щитовидного хряща прикрепляется к подъязычной кости с помощью тирео-подъязычной мембраны . Эта фиброэластичная плита имеет толстую срединную область, называемую медианной подъязычно-подъязычной связкой , а также боковые части, называемые латеральными подъязычно-подъязычными связками , которые непосредственно прикрепляются к верхнему рогу.Нижний рог имеет фасетку на своей медиальной поверхности, с помощью которой он сочленяется с заднебоковой поверхностью перстневидного хряща , образуя перстневидный сустав.
перстневидный хрящ представляет собой гиалиновый хрящ в форме кольца гораздо меньшего размера, расположенный непосредственно под щитовидным хрящом. Он образует нижнюю часть гортани и соединяется с трахеей снизу. Это единственное полное хрящевое кольцо, окружающее дыхательные пути. Перстневидный хрящ имеет узкую переднюю дугу (участок полосы) и более широкую заднюю пластинку (участок печати) со срединным гребнем, который служит поверхностью прикрепления для пищевода.Плотная связка соединительной ткани, перстневидная связка , прикрепляет перстневидный хрящ к нижнему краю щитовидного хряща. Крикотрахеальная связка прикрепляет перстневидный хрящ к первому кольцу трахеи.
На суполатеральных сторонах более широкой пластинки заднего перстневидного хряща расположены парные пирамидальные черпаловидные хрящи . Каждый хрящ имеет верхнюю вершину, передний голосовой отросток и большой латеральный мышечный отросток.К вершинам черпаловидного хряща прикреплены маленькие парные конические хрящи конической формы .
Последний непарный хрящ, надгортанник , представляет собой большой пластичный эластичный хрящ, покрытый слизистой оболочкой. Он прикрепляется своей ножкой к внутренней части угла, образованного пластинками щитовидного хряща, через щитовидно-надгортаную связку по средней линии. Надгортанник также прикреплен к подъязычной кости подъязычной связкой , которая проходит от переднегортативной поверхности надгортанника до тела подъязычной кости.Надгортанник выступает выше глотки, а его верхний край располагается сразу за корнем языка. Как следует из названия (epi = вверху, glottis = устье дыхательного горла), он находится над отверстием гортани (входным отверстием). Во время глотания, когда гортань движется вверх и вперед, надгортанник опускается вниз, закрывая входное отверстие в гортани и тем самым предотвращая попадание материалов в дыхательные пути.
Тонкий слой соединительной ткани, четырехугольная мембрана проходит между латеральными краями надгортанника и переднебоковыми краями черпаловидного хряща.Ее свободный нижний край утолщен и образует вестибулярную связку . Эта связка окружена складкой слизистой оболочки, образуя вестибулярную складку (ложный голосовой связок), которая простирается от хряща щитовидной железы к черпаловидному хрящу.
Последними хрящевыми компонентами гортани являются два небольших булавовидных клиновидных хрящей , которые лежат выше и кпереди от роговых хрящей. Они не прикрепляются напрямую к каким-либо другим хрящам гортани, но подвешены внутри и укрепляют фиброэластическую мембрану, называемую надгортанной мембраной .Эта мембрана образует свободный верхний край четырехугольной мембраны, которая, как описано ранее, соединяет черпаловидные хрящи с боковыми границами надгортанника. Он покрыт слизистой оболочкой, образуя надгортанниковую складку . На задней стороне надгортанника и роговой, и клиновидный хрящи видны как небольшие узелки, окружающие входное отверстие гортани.
Полость гортани
Теперь, когда мы обсудили внешний хрящевой скелет, давайте заглянем внутрь просвета гортани.Выложенная слизистой оболочкой полость гортани простирается от ее верхнего отверстия (входа в гортань) до нижней границы перстневидного хряща, которая продолжается с просветом трахеи. Полость гортани делится на три области:
- Преддверие : между входом в гортань и вестибулярными складками
- Средняя часть : между вестибулярными складками вверху и голосовыми складками внизу
- Подглоточная полость : между голосовой складкой и трахеей
Боковые стенки средней части полости гортани выступают наружу, образуя боковые выемки ( гортанный желудочек, ) между вестибулярной складкой и голосовой складкой.Каждый желудочек имеет удлиненное слепое трубчатое расширение, гортанный мешок , который выступает впереди вверх между вестибулярной складкой и щитовидным хрящом. Каждый мешочек выстлан слизистыми железами, которые вырабатывают слизь для смазывания голосовых складок.
Голосовой аппарат гортани называется голосовой щелью и состоит из двух голосовых складок (настоящие голосовые связки) и голосовых связок . Каждая голосовая складка, покрытая слизистой оболочкой, содержит голосовую связку, которая простирается от внутренней поверхности щитовидного хряща до голосового отростка соответствующего черпаловидного хряща.Параллельно каждой голосовой связке проходит голосовая мышца, отвечающая за регулировку напряжения голосовых связок. Голосовые связки — это «настоящие» структуры, которые производят звук при прохождении над ними воздуха, тогда как вестибулярные складки (ложные голосовые связки) не играют никакой роли в производстве звука, но защищают голосовые связки.
Отверстие между голосовыми связками называется голосовой связкой. Форма этого отверстия зависит от положения голосовых связок. Вращательные движения черпаловидных хрящей в перстневидных суставах могут разделять (отводить) голосовые складки, расширять голосовую щель или соединять (соединять) голосовые складки и сужать голосовую оболочку.Обычно во время дыхания голосовые связки отводятся, однако во время глотания они сводятся, чтобы закрыть голосовую щель. Качество производимого звука (например, высота звука) зависит от длины, напряжения и положения голосовых связок. Обычно во время звучания голосовые связки сближаются, образуя щелевидную голосовую щель. Когда выдыхаемый воздух проходит по шнурам, он заставляет их вибрировать, производя звук. Затем звук преобразуется движениями языка, челюсти и губ в речь.
Мышцы
С гортани связаны две группы мышц: внешние и внутренние мышцы. Внешние мышцы гортани перемещают гортань в целом. Они состоят из надподъязычных мышц, которые поднимают подъязычную кость и гортань во время глотания и вокализации, и подъязычных мышц, которые вдавливают подъязычную кость и гортань.
Маленькие внутренние мышцы гортани отвечают за движение различных компонентов гортани.Они изменяют длину и напряжение голосовых связок, а также форму голосовой щели во время дыхания, глотания и вокализации.
Мышцы гортани (обзорная схема)
Кровоснабжение и лимфодренаж
Артериальное кровоснабжение гортани осуществляется через верхнюю и нижнюю гортанные артерии , которые являются ветвями артерий щитовидной железы . Гортань дренируется соответствующими венами, а именно верхней и нижней гортанными венами.Лимфатические сосуды над голосовыми складками стекают в верхние глубокие шейные лимфатические узлы узлов , тогда как сосуды ниже голосовых складок стекают сначала в узлы вокруг трахеи (предтрахеальные и паратрахеальные узлы), а затем в нижние глубокие шейные лимфатические узлы.
Иннервация
Правый и левый верхние и нижние гортанные нервы , которые являются ветвями блуждающего нерва, десятого черепного нерва (CN X), обеспечивают моторную и сенсорную иннервацию гортани.Каждый верхний гортанный нерв делится на внутренних и внешних гортанных нервов . Внутренний гортанный нерв сопровождает верхнюю гортанную артерию через щитовидно-подъязычную перепонку и обеспечивает сенсорную и вегетативную иннервацию полости гортани до уровня голосовых связок. Наружный гортанный нерв меньшего размера обеспечивает двигательную иннервацию перстневидно-щитовидной мышцы.
возвратных гортанных нервов , которые являются восходящими ветвями блуждающих нервов, продолжаются по направлению к гортани как правый и левый нижние гортанные нервы.Они обеспечивают двигательную иннервацию всех внутренних мышц гортани, за исключением перстнещитовидных мышц , и сенсорную иннервацию полости гортани ниже голосовых связок.
Клиническое применение
При острой опасной для жизни обструкции дыхательных путей врачи могут выполнить крикотиротомию , вставив иглу через перстневидную связку для обеспечения проходимости дыхательных путей. Для выполнения этой процедуры требуется базовое понимание анатомии гортани.В стерильных условиях врачи пальпируют выступ гортани щитовидного хряща (кадык) и скользят пальцами вниз, пока не почувствуют перстневидный хрящ, который представляет собой первую ощутимую твердую выпуклость. Зазор между щитовидным и перстневидным хрящами (перстнещитовидное пространство , ) перекрывается перстнещитовидной связкой.
Интересный факт
Дети старшего возраста и взрослые не могут глотать и дышать носом одновременно. Однако новорожденные и младенцы могут делать это при глотании молока.Этот процесс называется « облигатное носовое дыхание ». У младенцев гортань расположена выше, и при дальнейшем подъеме во время глотания надгортанник может скользить вверх за мягкое небо, запирая гортань в носоглотке. Это анатомическое устройство, которое позволяет младенцам кормить и дышать одновременно, теряется в возрасте от 2 до 6 лет, поскольку гортань постепенно опускается во взрослое место.
Как исправить проблемы сопряжения Bluetooth
Bluetooth — это популярный метод беспроводной передачи данных между двумя устройствами, например телефоном и наушниками, медиаплеером и динамиком или iPad и клавиатурой.По словам Сукэ Джаванды, директора по маркетингу компании Bluetooth Special Interest Group, это одна из наиболее широко используемых беспроводных технологий в мире. Он говорит, что только в этом году будет отгружено около 2,5 миллиардов устройств Bluetooth, и это число, вероятно, удвоится в течение следующих четырех лет.
Bluetooth великолепен, когда он работает. Но если вы тот, кто любит поиграть с такими подключенными гаджетами, вы знаете, что может быть неприятно, когда между ними происходит зависание.Вот несколько распространенных причин проблем со спариванием, а также советы о том, что с ними делать.
Почему не удается установить пары Bluetooth
Bluetooth зависит от аппаратного и программного обеспечения для правильной работы. Такие производители, как Apple и Microsoft, все чаще включают радиостанции, говорящие на языке Bluetooth 4.0, в свое новое оборудование. Но если у вас есть устройство, например телефон двухлетней давности, в котором отсутствует этот новый вид радио, вы не сможете подключиться к устройствам, поддерживающим только Bluetooth 4.0.
Когда я недавно тестировал множество спортивных часов, мой Samsung Galaxy Nexus, оснащенный радиомодулем Bluetooth 3.0, не смог подключиться к Mio Alpha, инновационному устройству, которое отслеживает частоту сердечных сокращений без нагрудного ремня. Однако iPhone 5 моего мужа мог связываться с Mio Alpha, потому что каждый iPhone, начиная с iPhone 4S, поддерживает Bluetooth Smart.
Хорошая новость, говорит мне Джаванда, заключается в том, что большинство телефонов, выпущенных за последние 18 месяцев, имеют двойной радиомодуль, который обменивается данными с устройствами Bluetooth Smart Ready, а также с устройствами, оснащенными предыдущими версиями Bluetooth.Так что, если ваш текущий телефон не поддерживает Bluetooth Smart, возможно, следующий будет.
Но даже если бы в моем телефоне был соответствующий тип радио для Bluetooth 4.0, моя текущая операционная система, Android 4.2.2, не поддерживает Bluetooth Smart. Пользователи Android не смогут использовать Bluetooth 4.0, пока не получат Android 4.3 (при условии, что у них есть подходящее оборудование).
Устройства
также поставляются со специальными профилями Bluetooth. Если Bluetooth является общеязыковым соединением устройств, вы можете думать о профиле как о диалекте, связанном с определенным использованием.Например, вы, вероятно, не сможете соединить мышь и камеру, потому что камера не поддерживает профиль устройства с интерфейсом пользователя. Но если и мобильный телефон, и беспроводная гарнитура поддерживают профиль громкой связи, у вас должна быть возможность выполнить их сопряжение.
Однако, если сбой сопряжения вызван ошибкой пользователя, вы можете предпринять шаги, чтобы ваши устройства успешно взаимодействовали друг с другом.
Что можно сделать при сбое сопряжения
1.Определите, какой процесс сопряжения использует ваше устройство. Процесс сопряжения устройств может отличаться. Иногда, например, требуется ввести код в телефон. В других случаях вам нужно физически прикоснуться телефоном к устройству, с которым вы хотите установить сопряжение; например, Sony SmartWatch 2 использует NFC (связь ближнего радиуса действия) в сочетании с Bluetooth для сопряжения. Или, в случае Nike + FuelBand, вам нужно всего лишь удерживать кнопку на часах, чтобы подключить их к iPhone.
Если вы не знаете, как выполнить сопряжение устройства, обратитесь к его руководству пользователя; обычно вы можете найти его, выполнив поиск в Интернете.
2. Убедитесь, что Bluetooth включен. Вы должны увидеть маленький значок Bluetooth в верхней части экрана телефона. Если вы этого не сделаете, войдите в настройки, чтобы включить его.
3. Включите режим видимости. Допустим, вы хотите связать свой телефон с информационно-развлекательной системой автомобиля, чтобы вы могли звонить, отправлять текстовые сообщения и пользоваться навигацией в режиме громкой связи. Сначала зайдите в настройки своего телефона и нажмите Bluetooth; телефон будет виден в машине. Затем нажмите кнопки на информационно-развлекательной системе вашего автомобиля, обычно на рулевом колесе или центральной консоли, чтобы он начал поиск устройства.
Как только он найдет ваш телефон, автомобиль отобразит числовой код, который необходимо подтвердить или ввести на телефоне. После этого устройства должны быть сопряжены. Имейте в виду, что ваш телефон будет оставаться в режиме видимости только несколько минут; если вы потратите слишком много времени, вам придется начинать заново.
4. Выключите и снова включите устройства. Мягкий сброс иногда может решить проблему. С телефонами это проще всего сделать, войдя в режим полета и выйдя из него.
5.Удалите устройство с телефона и откройте его заново. Если ваш телефон видит устройство, но не получает от него данные, иногда помогает начать с нуля. В настройках iOS вы можете удалить устройство, нажав на его имя, а затем Забыть это устройство. В настройках Android коснитесь имени устройства и затем «Разорвите пару». После удаления устройства начните с шага 1 в этом списке.
6. Убедитесь, что устройства, которые вы хотите соединить, предназначены для подключения друг к другу. Будь то гарнитура, динамик, мышь, клавиатура, камера или что-то еще, ваше устройство имеет специальный профиль, в котором указано, к чему оно может подключаться.Если вы не уверены, обратитесь к руководству пользователя.
7. Загрузите драйвер. Если у вас возникли проблемы с подключением чего-либо к компьютеру, возможно, вам не хватает нужного драйвера. Самый простой способ выяснить это — выполнить поиск в Интернете по имени устройства, которое вы пытаетесь связать, со словом «драйвер».
8. Обновите прошивку оборудования. Некоторые автомобильные аудиосистемы в последнее время не поддерживают сопряжение с iPhone 5, например, потому что драйверы Bluetooth в этих системах не работают с Bluetooth 4.0. Если вы не знаете, как получить последнюю версию прошивки для вашего оборудования, обратитесь к производителю устройства.
9. Убедитесь, что два устройства находятся достаточно близко друг к другу. Хотя вы бы не подумали, что кто-то может попытаться соединить iPad с клавиатурой, если бы эти двое не сидели рядом друг с другом, вероятно, стоит отметить, что вы должны убедиться, что все устройства, которые вы пытаетесь соединить, находятся в пределах пяти ноги друг друга.
10. Помните, что не все беспроводные устройства используют Bluetooth.Альтернативы включают спецификацию Wireless Gigabit, Wireless HD, ANT +, ZigBee, NFC, а также Wi-Fi Direct. Эти другие технологии обычно не работают с вашим телефоном, планшетом или ПК, отмечает Джаванда, без какого-либо дополнительного оборудования.
Мы надеемся, что это руководство помогло вам решить проблемы с сопряжением Bluetooth. Если вы знаете какой-либо совет, который мы упустили, поделитесь в комментариях ниже!
Эта статья была написана Кристиной ДесМаре и первоначально появилась на Techlicious.
Больше от Techlicious:
.