
Содержание
Биология для студентов — 14. Темновая фаза фотосинтеза. Исследования Кальвина
Темновая фаза фотосинтеза связана с реакциями фиксации углерода, которые проходят в строме хлоропласта и продолжаются в цитоплазме без непосредственного поглощения света. В процессе световой фазы фотосинтеза накапливается достаточно высокий уровень АТФ и НАДФ·Н. Однако сами по себе эти макроэргические соединения не способны синтезировать углеводы из CO2. Становится очевидным, что и темновая фаза фотосинтеза – сложный процесс, включающий большое количество последовательно идущих реакций, возможных только после осуществления световой фазы.
Существует несколько разных путей связывания CO2 в углеводы, встречающихся у растений разных экологических и систематических групп, но основным, характерным для всех растений, является так называемый C3-путь фотосинтеза, или цикл Кальвина.
Способ ассимиляции СО2 в углеводы, присущий всем растениям, был расшифрован только в середине XX века американским биохимиком Мэлвином Кальвином и его коллегами на примере одноклеточных зеленых водорослей (хлореллы и др.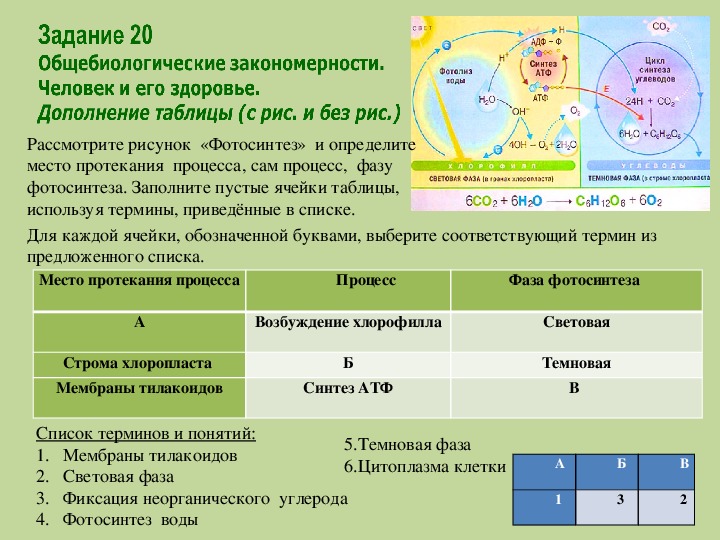 ) и зеленых листьев шпината. Исследование этой проблемы продолжалось 10 лет – с 1946 по 1966 год. Вначале ученые вели поиск первичного акцептора CO2. После ряда экспериментов они установили, что первичную фиксацию CO2 осуществляет пятиуглеродный сахар – рибулозо-1,5-дифосфат РуДФ). Фиксация осуществляется следующим образом: сначала происходит присоединение CO2 к молекуле РуДФ. При этом образуется промежуточный продукт – очень неустойчивое шестиуглеродное соединение, из которого в присутствии воды образуются две молекулы трехуглеродного соединения – 3-фосфоглицериновой кислоты (3-ФГК). В этой реакции для связывания одной молекулы CO2 затрачивается три молекулы АТФ и две молекулы НАДФ·Н.
) и зеленых листьев шпината. Исследование этой проблемы продолжалось 10 лет – с 1946 по 1966 год. Вначале ученые вели поиск первичного акцептора CO2. После ряда экспериментов они установили, что первичную фиксацию CO2 осуществляет пятиуглеродный сахар – рибулозо-1,5-дифосфат РуДФ). Фиксация осуществляется следующим образом: сначала происходит присоединение CO2 к молекуле РуДФ. При этом образуется промежуточный продукт – очень неустойчивое шестиуглеродное соединение, из которого в присутствии воды образуются две молекулы трехуглеродного соединения – 3-фосфоглицериновой кислоты (3-ФГК). В этой реакции для связывания одной молекулы CO2 затрачивается три молекулы АТФ и две молекулы НАДФ·Н.
Схема первичной фиксации CO2
Реакцию фиксации углерода, открытую в 1948 году, катализирует очень крупный фермент из стромы хлоропласта – рибулозобисфосфаткарбоксилазаоксигеназа (сокращенно – РУБИСКО). Так как фермент РУБИСКО работает весьма медленно, необходимо, чтобы его молекул в хлоропластах было много.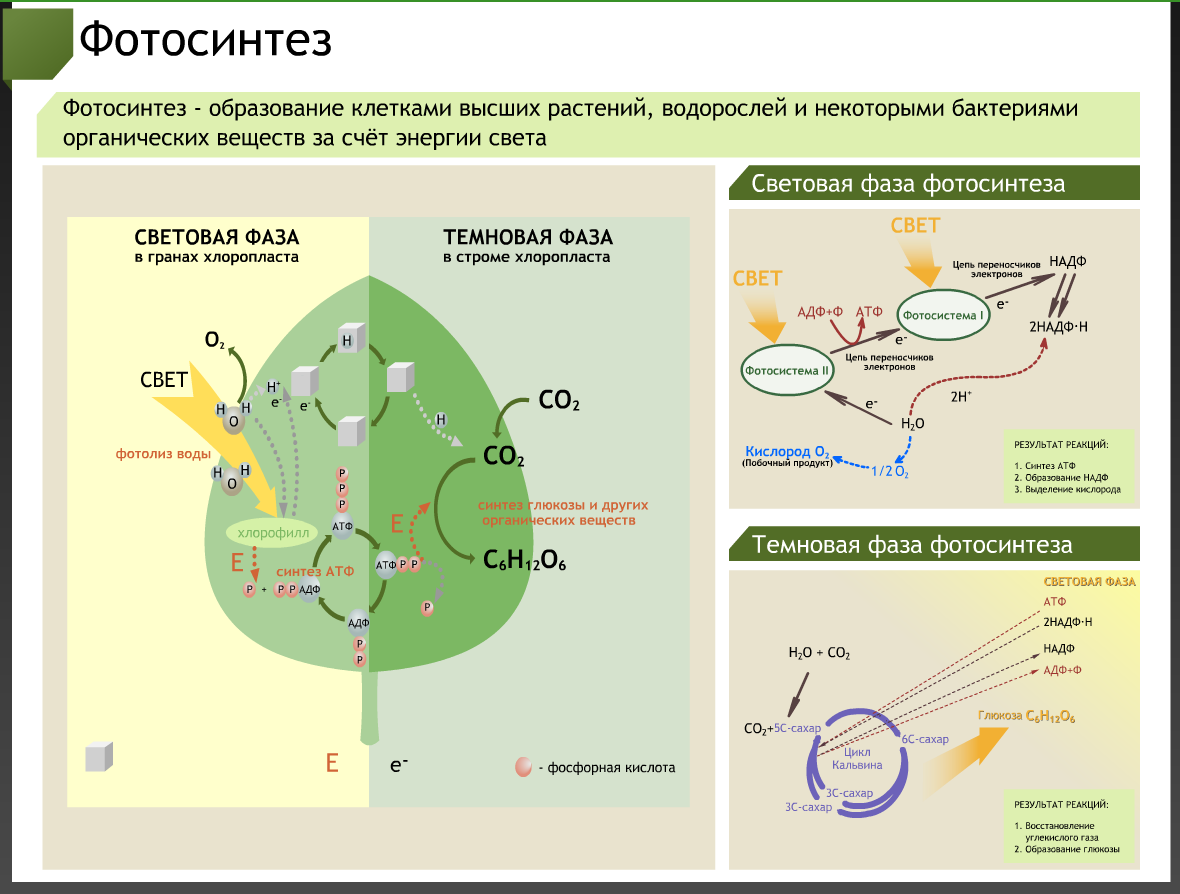 Действительно, этот фермент обычно составляет более 50 % общего количества белков хлоропластов. Многие исследователи утверждают, что это самый распространенный белок в живой природе.
Действительно, этот фермент обычно составляет более 50 % общего количества белков хлоропластов. Многие исследователи утверждают, что это самый распространенный белок в живой природе.
Дальнейшие исследования лаборатории Кальвина способствовали установлению всех последующих реакций C3-пути фотосинтеза, обеспечивающих синтез углеводов. За расшифровку механизма фиксации CO2 в процессе фотосинтеза М. Кальвин в 1961 году стал лауреатом Нобелевской премии по химии.
Цикл Кальвина состоит из трех стадий:
- карбоксилирования,
- восстановления,
- превращения.
Упрощенная схема цикла Кальвина – пути фиксации углерода при фотосинтезе
На первой стадии (карбоксилирование) фиксация углерода идет с участием ферментов и АТФ, полученной на световой фазе фотосинтеза; при этом образуются молекулы 3-фосфоглицериновой кислоты (3-ФГК). На второй стадии (восстановление) помимо АТФ используется НАДФ·Н. Здесь 3-ФГК восстанавливается до 3-фосфоглицеринового альдегида (З-ФГА), часть молекул которого идет на синтез 6-углеродного моносахарида (глюкозы или фруктозы).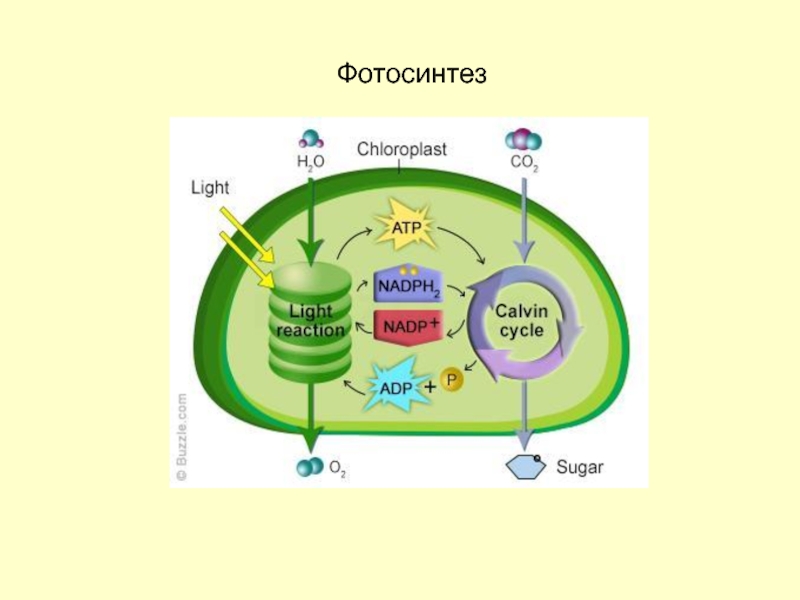 На третьей стадии (превращение) при повторении цикла другая часть молекул 3-ФГА используется для синтеза шестиатомного фосфорилированного моносахарида – фруктозо-1,5-дифосфата. Трехуглеродные фосфосахара вместе с множеством других продуктов метаболизма хлоропластов транспортируются в цитоплазму клетки, где образуют ди- и полисахариды (сахара, крахмал, целлюлозу или другие соединения).
На третьей стадии (превращение) при повторении цикла другая часть молекул 3-ФГА используется для синтеза шестиатомного фосфорилированного моносахарида – фруктозо-1,5-дифосфата. Трехуглеродные фосфосахара вместе с множеством других продуктов метаболизма хлоропластов транспортируются в цитоплазму клетки, где образуют ди- и полисахариды (сахара, крахмал, целлюлозу или другие соединения).
В процессах темновой фазы фотосинтеза образуются углеводы – первичные органические вещества. На определенном этапе темновой фазы фотосинтеза судьба трехуглеродных молекул 3-фосфоглицериновой кислоты может оказаться различной. Одни из них соединяются друг с другом и образуют шестиуглеродные сахара, которые, в свою очередь, могут полимеризоваться в крахмал, целлюлозу и др. Некоторые могут использоваться для синтеза аминокислот, карбоновых кислот, спиртов и пр. Но целый ряд молекул ФГК вовлекается в длинный ряд реакций, приводящих к превращению трехуглеродных молекул в молекулы пятиуглеродного сахара (РуДФ), которые могут снова ассимилировать углекислый газ и многократно повторять этот цикл до тех пор, пока растение живет и получает световую энергию.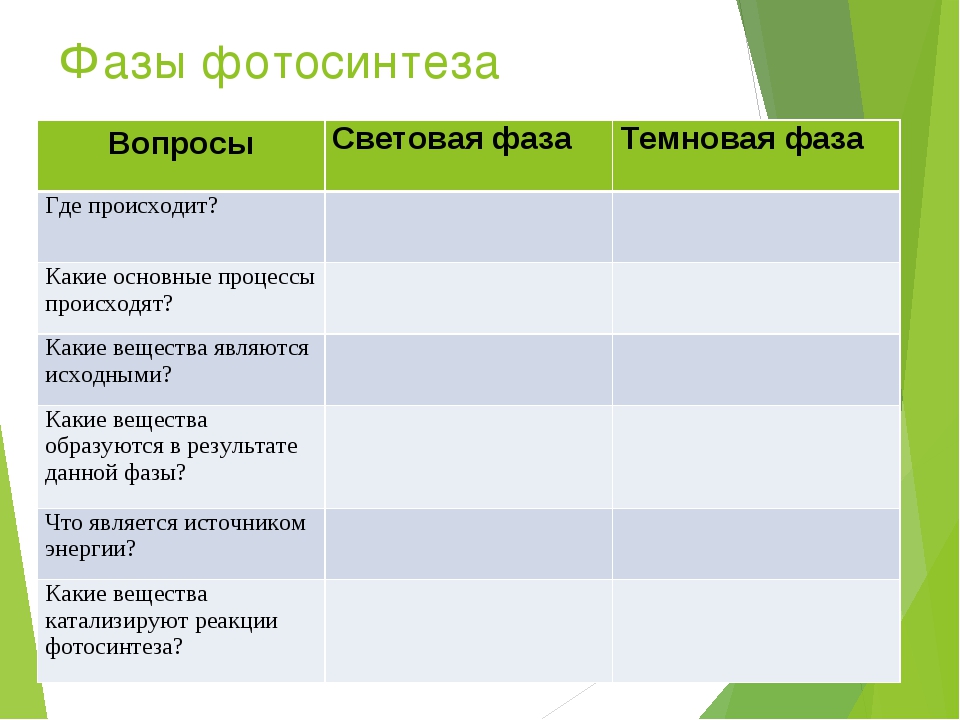 Все процессы темновой фазы фотосинтеза идут без непосредственного потребления света, но в них большую роль играют макроэргические соединения (АТФ и НАДФ·Н), образующиеся во время световой фазы фотосинтеза. Доказано, что для синтеза одной молекулы глюкозы в цикле Кальвина необходимы 12 молекул НАДФ·Н и 18 молекул АТФ, которые поставляются с тилакоидных мембран в результате фотохимических реакций световой фазы фотосинтеза.
Все процессы темновой фазы фотосинтеза идут без непосредственного потребления света, но в них большую роль играют макроэргические соединения (АТФ и НАДФ·Н), образующиеся во время световой фазы фотосинтеза. Доказано, что для синтеза одной молекулы глюкозы в цикле Кальвина необходимы 12 молекул НАДФ·Н и 18 молекул АТФ, которые поставляются с тилакоидных мембран в результате фотохимических реакций световой фазы фотосинтеза.
Схема синтеза 3-фосфоглицеринового альдегида (ФГА) в хлоропласте
В процессе темновой фазы энергия макроэргических связей АТФ преобразуется в химическую энергию органических соединений – молекул углеводов. Это означает, что энергия солнечного света как бы консервируется в химических связях между атомами органических веществ, что имеет огромное значение для энергетики биосферы и жизнедеятельности всего населения нашей планеты.
В настоящее время известны и другие пути ассимиляции углекислого газа наряду с системой его фиксации в цикле Кальвина (C3-пути фотосинтеза). Существует так называемый C4-путь ассимиляции углерода в фотосинтезе. Он может протекать при низких концентрациях CO2. Этот тип фиксации углекислого газа в фотосинтезе выработался в процессе эволюции у растений жарких, засушливых мест и наблюдается у кукурузы, сахарного тростника, проса, сорго, амаранта, лебеды, баклажанов и др., а также у растений, устойчивых к засолению почвы.
Существует так называемый C4-путь ассимиляции углерода в фотосинтезе. Он может протекать при низких концентрациях CO2. Этот тип фиксации углекислого газа в фотосинтезе выработался в процессе эволюции у растений жарких, засушливых мест и наблюдается у кукурузы, сахарного тростника, проса, сорго, амаранта, лебеды, баклажанов и др., а также у растений, устойчивых к засолению почвы.
Существует и особый тип фотосинтеза у таких растений, как кактусы, молочаи, крассулы, каланхое, седумы и другие суккуленты, произрастающие в засушливых, безводных условиях. Эти растения запасают CO2 в виде органических кислот ночью, так как он поступает в клетки только тогда, когда открыты их устьица (днем они закрыты для предотвращения потери воды).
Фотосинтез. Световая фаза.
Прежде чем перейти непосредственно к рассмотрению фотосинтеза, на всякий случай напомним немного про органеллы, в которых происходит большинство реакций, связанных с этим процессом – про хлоропласты.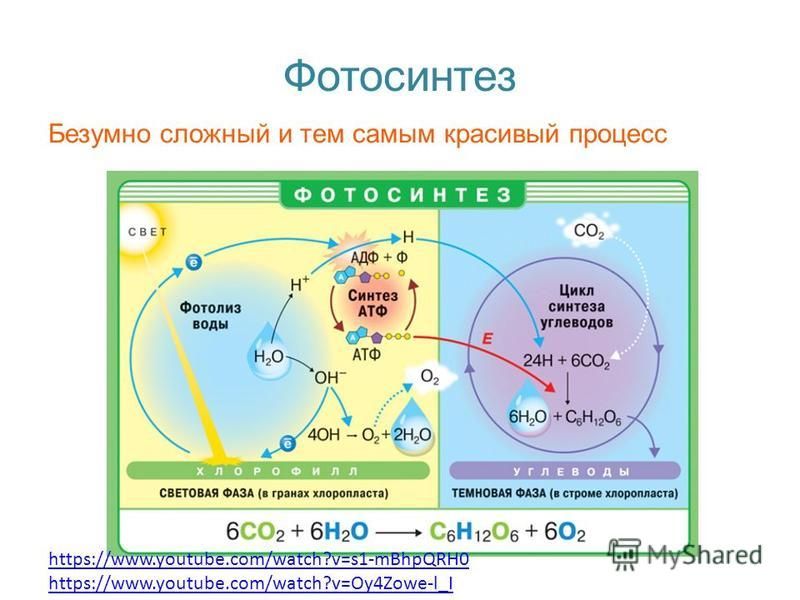 Пластиды были приобретены предком Архепластид как минимум 1,2 млрд лет назад путем эндосимбиоза с цианобактерией (см приложения в конце статьи). Архепластиды — супергруппа в современной систематике, включающая красных, зеленых, глаукофитовых, криптофитовых (Adl et al., 2018) водорослей, группу Streptophyta (харовые в широком смысле и наземные растения) и прочие группы.
Пластиды были приобретены предком Архепластид как минимум 1,2 млрд лет назад путем эндосимбиоза с цианобактерией (см приложения в конце статьи). Архепластиды — супергруппа в современной систематике, включающая красных, зеленых, глаукофитовых, криптофитовых (Adl et al., 2018) водорослей, группу Streptophyta (харовые в широком смысле и наземные растения) и прочие группы.
Хлоропласт, как и все пластиды – двумембранная органелла клетки. Наружная мембрана имеет происхождение из наружной мембраны цианобактерий, она проницаема для многих низкомолекулярных органических соединений. В наружной мембране содержится большое количество рецепторов и транслокаторов, за счет которых осуществляется обмен с внешней средой.
Внутренняя мембрана, о которой речь будет идти и далее, обладает высокой избирательностью проницаемости, контролирует транспорт, является основным барьером, разделяющим внутреннее содержимое пластиды от цитоплазмы и разграничивающим различные компартменты самого хлоропласта.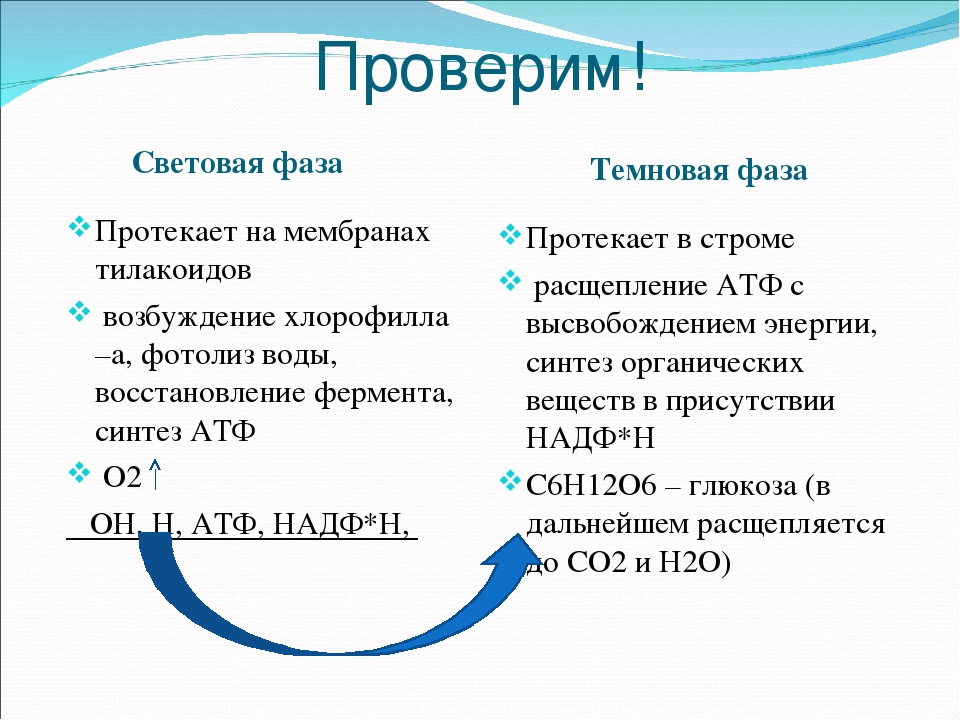 По происхождению она соответствует цитоплазматической мембране цианобактерии. Внутренняя мембрана образует обширную мембранную сеть внутри хлоропласта, образуя тилакоиды (небольшие мембранные пузырьки), часто собранные в стопки или граны и крупные свободные ламеллы стромы. Внутреннее пространство тилакоида и ламеллы называется люменом.
По происхождению она соответствует цитоплазматической мембране цианобактерии. Внутренняя мембрана образует обширную мембранную сеть внутри хлоропласта, образуя тилакоиды (небольшие мембранные пузырьки), часто собранные в стопки или граны и крупные свободные ламеллы стромы. Внутреннее пространство тилакоида и ламеллы называется люменом.
В высших растениях может быть большое разнообразие различных пластид, но в одной клетке только один тип. При этом многие типы пластид могут обратимо друг в друга превращаться.
Фотосинтез – один из основополагающих процессов биосферы, это наиболее мощный механизм преобразования солнечной энергии в энергию химических связей, что делает его роль ключевой в биоэнергетике как растений, так и животных.
Реакции фотосинтеза традиционно делят на две стадии – темновую и световую. И тут важно понимать, что, несмотря на название, обе стадии идут только на свету! Это объясняется тем, что многие ферменты, катализирующие ключевые реакции, активны только на свету. Более того, для многих из них обнаружена зависимость от циркадных ритмов, внутренних часов организма, определяющих суточную ритмику различных процессов.
Более того, для многих из них обнаружена зависимость от циркадных ритмов, внутренних часов организма, определяющих суточную ритмику различных процессов.
Световая фаза фотосинтеза включает в себя разнообразные процессы, основная цель которых – преобразование солнечной энергии в энергию химических связей. Эти процессы локализованы на мембране тилакоида. По сути люмен тилакоида аналогичен межмембранному пространству митохондрии. Как и внутренняя мембрана митохондрии оболочка тилакоида представляет собой сопрягающую мембрану. Сопрягаются и в том и другом случае процессы перекачки протонов и перенос электронов по электрон-транспортной цепи. Далее за счет градиента протонов синтезируется АТФ, а электроны восстанавливают NADP+ до NAPDH+H+.
Вообще, в клетках есть две основные формы энергии – энергия химических связей (АТФ, например) и энергия градиентов (градиент протонов у растений, натрия – у животных), с которой неразрывно связана разность потенциалов и для создания которой необходима мембрана с жестко контролируемой проницаемостью.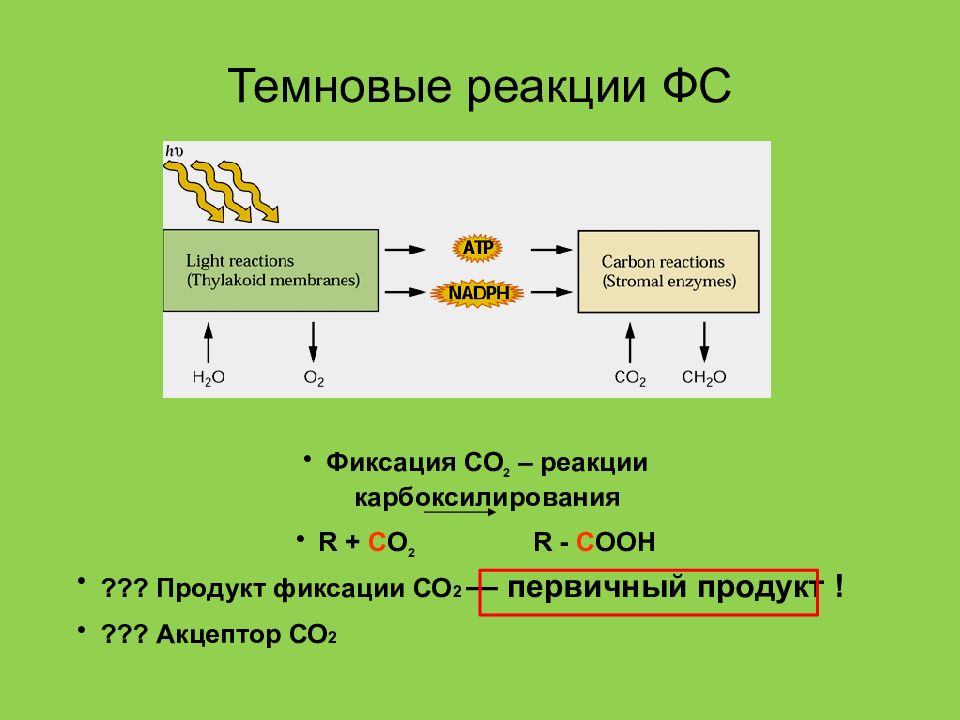 Не очень корректно, но использование энергии градиента можно сравнить с принципом работы гидроэлектростанций, где за счет разницы давления воды раскручиваются турбины и вырабатывается электричество. Эти две формы энергии обязательно присутствуют в любой живой клетке, а также могут быть конвертированны одна в другую. На этом факте основаны многие задания на олимпиадах, которые могут звучать как-то так:
Не очень корректно, но использование энергии градиента можно сравнить с принципом работы гидроэлектростанций, где за счет разницы давления воды раскручиваются турбины и вырабатывается электричество. Эти две формы энергии обязательно присутствуют в любой живой клетке, а также могут быть конвертированны одна в другую. На этом факте основаны многие задания на олимпиадах, которые могут звучать как-то так:
Рассчитайте сколько молекул АТФ может синтезироваться в митохондрии в аэробных условиях при полном окислении сахарозы?
Электронно транспортная цепь со сложным механизмом сопряжения возникла не сразу. Но рассмотрим все по порядку и начнем с предыстории.
Основными пигментами, участвующими в реакциях световой фазы являются хлорофиллы у эукариотических фотосинтезирующих организмов или бактериохлорофиллы у прокариотических, но самый первый механизм преобразования энергии света не задействовал эти пигменты. Речь про родопсиновый фотосинтез, встречаемый у группы архей Euryarchaeota (Halobacteria). Этот тип примечателен своим потрясающе простым устройством. Ключевым ферментом является собственно пигмент-белковый комплекс – бактериородопсин, в котором содержится ретиналь. Да, именно тот самый, что и в наших зрительных рецепторах. Под действием света ретиналь претерпевает цис-транс переход, что приводит к конформационным изменениям бактериородопсина и переносу протона из цитоплазмы наружу. Таким образом создается протонный градиент, который далее используется для синтеза АТФ на АТФ-синтазе (в данном случае не обычная, а архейного типа, похожая на вакуолярную АТФ-синтазу V-типа у растений). Прекрасный механизм, ничего лишнего!
Этот тип примечателен своим потрясающе простым устройством. Ключевым ферментом является собственно пигмент-белковый комплекс – бактериородопсин, в котором содержится ретиналь. Да, именно тот самый, что и в наших зрительных рецепторах. Под действием света ретиналь претерпевает цис-транс переход, что приводит к конформационным изменениям бактериородопсина и переносу протона из цитоплазмы наружу. Таким образом создается протонный градиент, который далее используется для синтеза АТФ на АТФ-синтазе (в данном случае не обычная, а архейного типа, похожая на вакуолярную АТФ-синтазу V-типа у растений). Прекрасный механизм, ничего лишнего!
Хлорофилльный фотосинтез появился позднее. Несмотря на все разнообразие хлорофиллов, только хлорофилл а и бактериохлорофилл а, располагаясь в димерах, могут непосредственно преобразовывать энергию света.
Все хлорофиллы имеют тетрапиррольную структуру порфирина, которая преимущественно и определяет свойства молекулы. Молекула плоская, несет систему сопряженных двойных связей, формирующих π-электронное облако, благодаря чему может довольно легко вступать в процессы фотоокисления (отдавать электрон) или фотовосстановления (принимать электрон). В центре хелатирован магний, который сильно влияет на свойства молекулы – выравнивает и перераспределяет электронную плотность по кольцам порфирина, образует координационные связи с молекулами воды, аминокислотными остатками, участвует в формировании димерных структур реакционных центров фотосистем. Также имеются полярные периферические группы, которые определяют взаимодействия хлорофиллов с различными компонентами хлоропласта. У многих хлорофиллов помимо тетрапиррольного кольца есть еще фитольный хвост. Фитол – дитерпеноид, содержащий 20 атомов углерода. Фитол практически не влияет на спектральные свойства хлорофилла, но определяет его взаимодействие с гидрофобной фракцией мембран и белков.
В центре хелатирован магний, который сильно влияет на свойства молекулы – выравнивает и перераспределяет электронную плотность по кольцам порфирина, образует координационные связи с молекулами воды, аминокислотными остатками, участвует в формировании димерных структур реакционных центров фотосистем. Также имеются полярные периферические группы, которые определяют взаимодействия хлорофиллов с различными компонентами хлоропласта. У многих хлорофиллов помимо тетрапиррольного кольца есть еще фитольный хвост. Фитол – дитерпеноид, содержащий 20 атомов углерода. Фитол практически не влияет на спектральные свойства хлорофилла, но определяет его взаимодействие с гидрофобной фракцией мембран и белков.
Синтез хлорофиллов идет в хлоропластах из глутамата, также как и синтез гемов (часть биосинтетического пути общая). Сначала происходят преобразования глутамата, получается соединение с пятичленным гетероциклом – порфобилиноген. Четыре порфобилиногена соединяются, получившаяся молекула претерпевает несколько превращений и получается протопорфирин IX.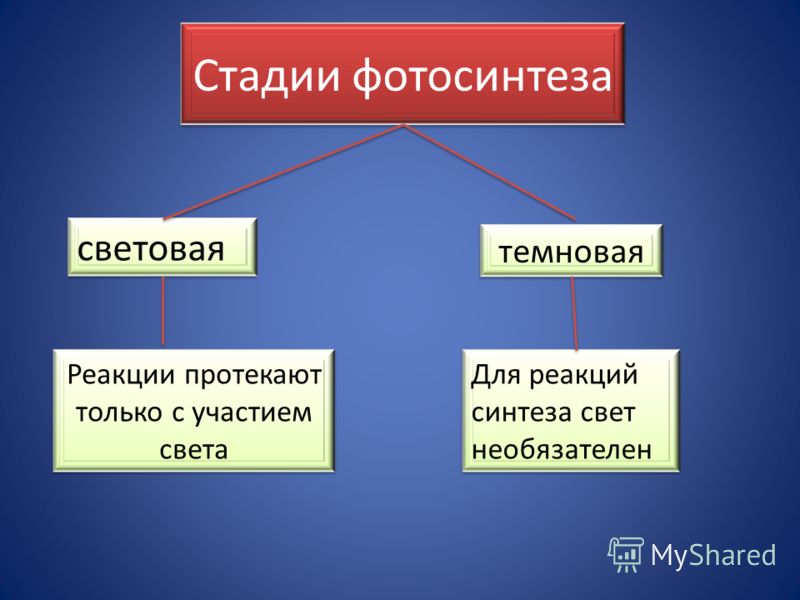 На этом соединении и образуется развилка в синтезах гемов и хлорофиллов. Если в соединение включается Mg2+, образуется Mg-протопорфирин и далее реакции идут по направлению биосинтеза хлорофилла. В темноте синтез хлорофиллов не может быть завершен, так как и ферменты контролируются светом и есть реакция, которая в принципе не идет в темноте. Последним этапом синтеза хлорофилл-синтаза присоединяет к тетрапирролу фитольный хвост, получается хлорофилл а. Все остальные хлорофиллы синтезируются из него.
На этом соединении и образуется развилка в синтезах гемов и хлорофиллов. Если в соединение включается Mg2+, образуется Mg-протопорфирин и далее реакции идут по направлению биосинтеза хлорофилла. В темноте синтез хлорофиллов не может быть завершен, так как и ферменты контролируются светом и есть реакция, которая в принципе не идет в темноте. Последним этапом синтеза хлорофилл-синтаза присоединяет к тетрапирролу фитольный хвост, получается хлорофилл а. Все остальные хлорофиллы синтезируются из него.
Хлорофилл а является основным фотосинтетическим пигментом эукариотических фотосинтезирующих организмов, цианобактерий (у которых это единственная форма хлорофилла). Максимумы поглощения приблизительно 430 и 670 нм, то есть в синей и красной области. Между этими максимумами «зеленый провал», в котором хлорофиллы практически не поглощают.
Хлорофилл b – дополнительный пигмент, характерный для высших растений, выполняет в основном светособирающую роль. Красноволновый максимум поглощения несколько сдвинут в коротковолновую область, благодаря чему «зеленый провал» несколько сужается.
Разнообразные формы хлорофилла с обнаруживаются у Dinophyta и Stramenopiles. У этого типа пигментов нет фитольного хвоста.
Хлорофилл d, f – цианобактериальные. В некоторой литературе можно найти, что хлорофилл d есть у красных водорослей, но это ошибка.
Хлорофилл e – по некоторым довольно старым источникам, имеется у одного лишь представителя Xanthophyceae.
Бактериохлорофиллов тоже много (a, b, c, d, e, f, g), основной – бактериохлорофилл a. Для бактериохлорофиллов характерны длинноволновые максимумы поглощения.
Тетрапиррольные пигменты могут быть не только циклические, но и линейные, называемые фикобилинами. Различают фикоэритрин, фикоцианин и аллофикоцианин (по увеличению длины волны максимумов поглощения). Фикобилины есть, например, у цианобактерий, красных и криптофитовых водорослей, где они выполняют роль светособирающих антенн. У цианобактерий и красных водорослей — фикобилипротеины.
Помимо хлорофиллов в пигментных системах в обязательном порядке присутствуют каротиноиды – кислород-содержащие (ксантофиллы) и бескислородные (каротины).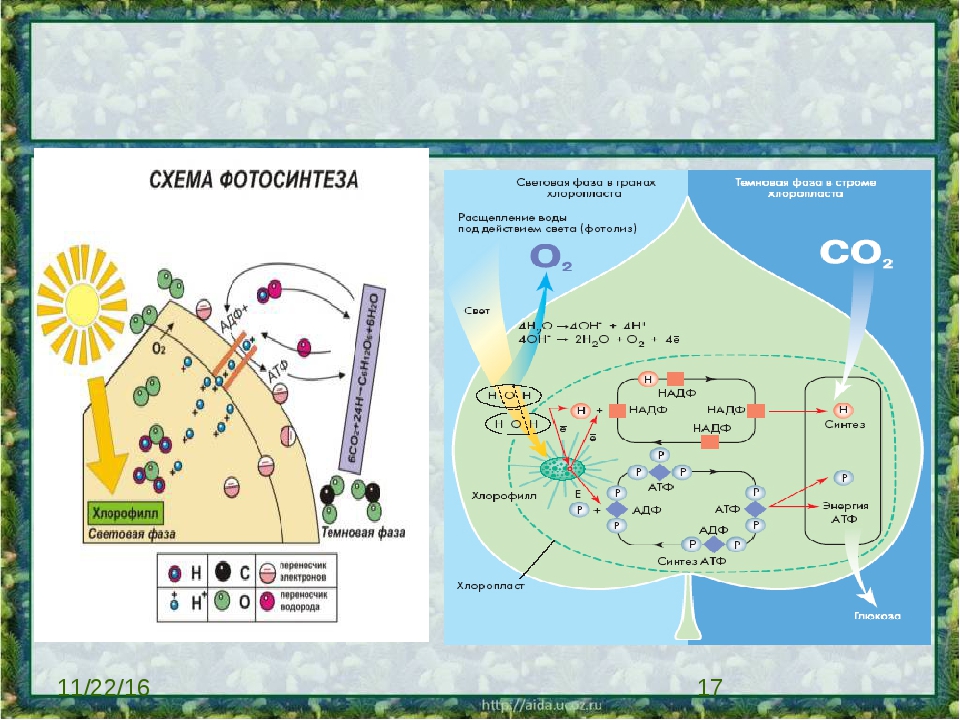 Это изопреноидные соединения с 40 атомами углерода, биосинтез которых происходит в пластидах. Благодаря длинной цепи сопряженных связей каротиноиды способны поглощать свет, передавать энергию возбуждения хлорофиллам, а также диссипировать ее в тепло. Этим частично объясняются фотопротекторные функции этих пигментов. Каротиноиды также могут забирать от хлорофиллов избыток энергии, тушить триплетный хлорофилл, защищать от активных форм кислорода (синглетный кислород, супероксидрадикал) и органических радикалов. Помимо этого, каротиноиды участвуют в структуризации мембран, определяют конформацию пигмент-белковых комплексов, придают окраску различным частям растений.
Это изопреноидные соединения с 40 атомами углерода, биосинтез которых происходит в пластидах. Благодаря длинной цепи сопряженных связей каротиноиды способны поглощать свет, передавать энергию возбуждения хлорофиллам, а также диссипировать ее в тепло. Этим частично объясняются фотопротекторные функции этих пигментов. Каротиноиды также могут забирать от хлорофиллов избыток энергии, тушить триплетный хлорофилл, защищать от активных форм кислорода (синглетный кислород, супероксидрадикал) и органических радикалов. Помимо этого, каротиноиды участвуют в структуризации мембран, определяют конформацию пигмент-белковых комплексов, придают окраску различным частям растений.
Что же все-таки такое антенна? Антенна – совокупность пигмент-белковых комплексов, способная улавливать свет и передавать энергию к реакционному центру.
Основная функция – увеличить площадь сбора световой энергии. В антеннах пигменты (хлорофиллы и каротиноиды) очень тесно друг с другом взаимодействуют, что позволяет им обмениваться энергией.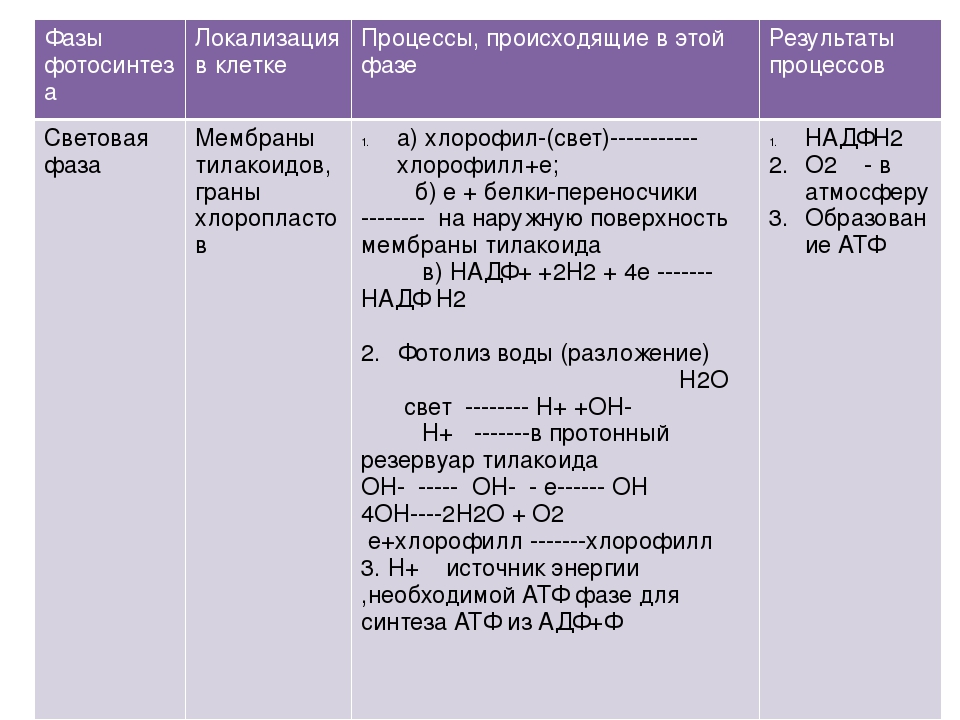 Говоря очень упрощенно, по антеннам перемещается экситон – некоторое возбужденное состояние, которое передается от пигмента с большей энергией возбуждения к пигменту с меньшей, попадая в конечном счете на реакционный центр, который таким образом выступает в качестве энергетической ловушки. Все процессы происходят очень быстро – за пикосекунды. Если говорить чуть подробнее, то в антеннах между хлорофиллами энергия передается путем образования переменного электрического поля между двумя пигментами, предлагается механизм осциллирующего диполя. Можно это представить на некотором механическом аналоге – в виде связанных колебаний.
Говоря очень упрощенно, по антеннам перемещается экситон – некоторое возбужденное состояние, которое передается от пигмента с большей энергией возбуждения к пигменту с меньшей, попадая в конечном счете на реакционный центр, который таким образом выступает в качестве энергетической ловушки. Все процессы происходят очень быстро – за пикосекунды. Если говорить чуть подробнее, то в антеннах между хлорофиллами энергия передается путем образования переменного электрического поля между двумя пигментами, предлагается механизм осциллирующего диполя. Можно это представить на некотором механическом аналоге – в виде связанных колебаний.
Представьте два шарика на упругих нерастяжимых подвесах, соединенных неким передаточным механизмом. Если начать периодически колебать один шарик, то со временем колебания передадутся на другой. На рисунке 9 схематично изображены процессы происходящие при этом на электронных орбиталях молекул.
Миграция энергии от каротиноида к хлорофиллу может идти и иным механизмом, что связано с особенностями возбуждения каротинов и стабильностью их возбужденных состояний.
Таким образом, в антеннах передается именно энергия, а все электроны остаются связанными со своими молекулами. В реакционном центре же под действием этой энергии происходит разделение зарядов в димере хлорофилла а и выбитый электрон поступает в электрон-транспортную цепь, на акцептор (см рисунок 7). А образовавшаяся «дырка» на пигменте реакционного центра восполняется за счет притока электронов от донора.
Существует несколько механизмов, каким образом восполнить ушедший электрон, в качестве доноров могут выступать различные соединения. Например, сероводород у анаэробных фотосинтезирующих зеленых серных бактерий, вода у эукариот и цианобактерий или пул хинолов, который в свою очередь принял электроны от фотосистемы. Но обо всем по порядку.
Электрон-транспортная цепь (ЭТЦ) хлоропласт высших растений состоит из двух фотосистем (ФС I и ФС II), цитохромного комплекса (цитохром b6/f), пула хинонов, пластоцианина, светособирающих антенн и некоторых стромальных белков.
В ЭТЦ есть несколько типов переносчиков.
Хиноны (окисленная форма, восстановленная – хинол) – гидрофобные двуэлектронные, располагаются внутри мембраны. Могут находиться в трех состояниях окисленности-восстановленности (редокс статус). Переносят электроны со второй фотосистемы на цитохромный комплекс. Помимо электронов переносят также и протоны, благодаря чему формируется протонный градиент.
Пластоцианин – медь-содержащий одноэлектронный белковый переносчик, располагающийся в люмене тилакоида.
Гемы – одноэлектронные переносчики, кофакторы цитохромов.
Железосерные кластеры – комплексы, состоящие из элементов железа и серы. Различаются по геометрии, числу и соотношению железа и серы. Непосредственно связаны с аминокислотами белков – цистеином и гистидином чаще всего. Переносят один (Fe-S, 2Fe-2S) или 2 электрона (4Fe-4S) за счет изменения степени окисленности железа. Железосерные кластеры имеются в цитохромном комплексе (белок Риске), в первой фотосистеме и в ферредоксине. Ферредоксин – стромальный белок, принимающий электроны от первой фотосистемы и передающий их на NADP+.
Прежде чем поговорить об ЭТЦ высших растений, необходимо сперва упомянуть прототипы ее компонентов.
Вторая фотосистема берет свое начало от фотосистемы пурпурных бактерий.
Система димерная (на рисунке 13 не показано), состоит из двух очень похожих белков (L и M), которые несут в себе одинаковые по переносчикам транспортные цепи (бактериохлорофилл-бактериофеофитин-хинон), но работает только цепь на L белке. Еще существует крупная субъединица H, которая защищает ЭТЦ внутри фотосистемы от внешних акцепторов. Пигментом в реакционном центре здесь является P870. Первичный акцептор электронов – бактериофеофитин (хлорофилл без магния, но с двумя протонами вместо него). Здесь же участвует бактериохлорофилл (BChl800). С бактериофеофитина электрон поступает на хиноны. QA (QLMQ) – связанный хинон, QB (QMPQ) – свободный, выходит в мембрану и несет два электрона к цитохромному комплексу, где окисляется, отдавая электроны и выносит два протона в люмен. Тут не b6/f цитохром, а bc1, как в митохондриях.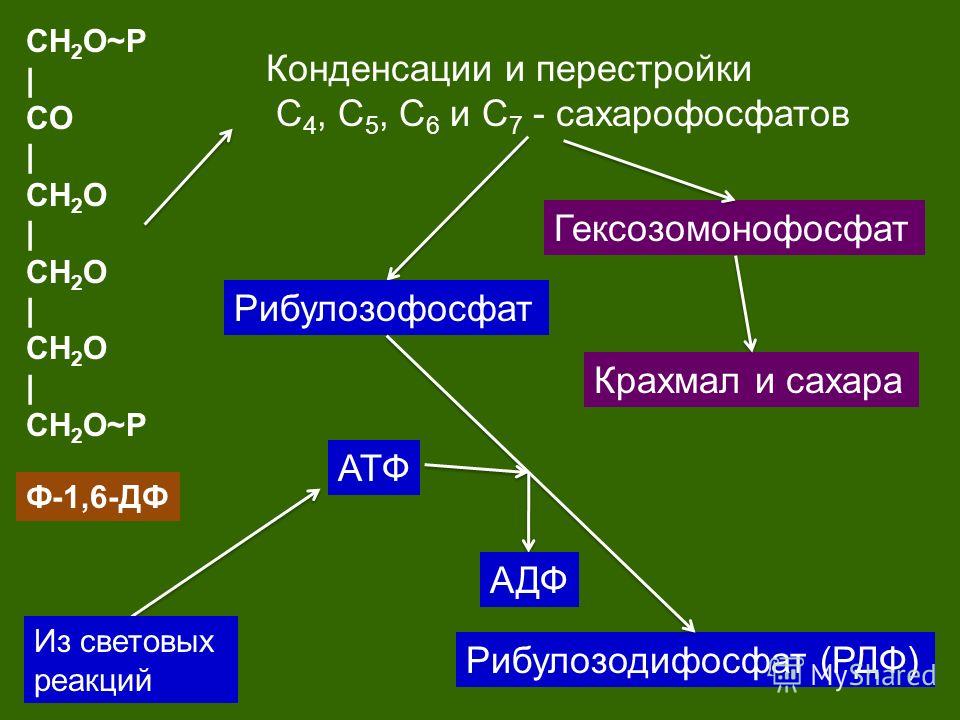 С цитохрмного комплекса электроны поступают на подвижный люменальный цитохром с, который возвращает их на реакционный центр.
С цитохрмного комплекса электроны поступают на подвижный люменальный цитохром с, который возвращает их на реакционный центр.
Этот тип фотосинтеза позволяет накачать протоны и синтезировать АТФ но не дает восстановительного эквивалента (NADPH), который пурпурным бактериям приходится образовывать, используя обратный транспорт электронов по ЭТЦ. Но это уже совсем другая история.
Фотосистема зеленых серных бактерий является прообразом первой фотосистемы эукариот и цианобактерий.
Пигмент в данном случае – Р840. Первичный акцептор – бактериохлорофилл A0, далее электрон идет на филлохинон А1 (витамин К1) и на железосерные кластеры Fx, FA, FB, с них на ферредоксин (ФД). Ферредоксин взаимодействует с фд-NADPH-оксидоредуктазой которая восстанавливает NADPH электронами с ФД. В данном случае электрон израсходован на образование восстановительного эквивалента, но надо также, как и в предыдущем случае, заполнить образовавшуюся «дырку» на пигменте реакционного центра. У зеленых серных фотосинтезирующих бактерий донором является сероводород, который, отдавая электроны, окисляется до серы или сульфатов. Отобранные от h3S электроны поступают через хинолы на цитохромный комплекс (снова bc1), а затем переносятся цитохромом с на реакционный центр. В данном случае, в противоположность тому, что имеется у пурпурных, идет образование NADPH, но не АТФ. Позволить себе получать сразу и NADPH и АТФ могут только эукариоты и цианобактерии, которые, вообще говоря, и дали начало пластидам.
Отобранные от h3S электроны поступают через хинолы на цитохромный комплекс (снова bc1), а затем переносятся цитохромом с на реакционный центр. В данном случае, в противоположность тому, что имеется у пурпурных, идет образование NADPH, но не АТФ. Позволить себе получать сразу и NADPH и АТФ могут только эукариоты и цианобактерии, которые, вообще говоря, и дали начало пластидам.
В хлоропластах и цианобактериях ЭТЦ представляет собой комбинирование двух описанных бактериальных систем (рисунок 15).
Окислительно-восстановительный (редокс) потенциал – способность химического вещества присоединять электроны. Выражается в милливольтах. Чем более отрицательные значения принимает редокс потенциал, тем лучшим донором электронов является вещество, чем более положительный – более сильным акцептором. Обратите внимание, что на схемах (рисунки 13-16) ось ординат отображает восстановительную способность, поэтому отрицательные значения сверху.
На рисунках 16 и 17 схематично показана общая логика процессов, происходящих в ЭТЦ.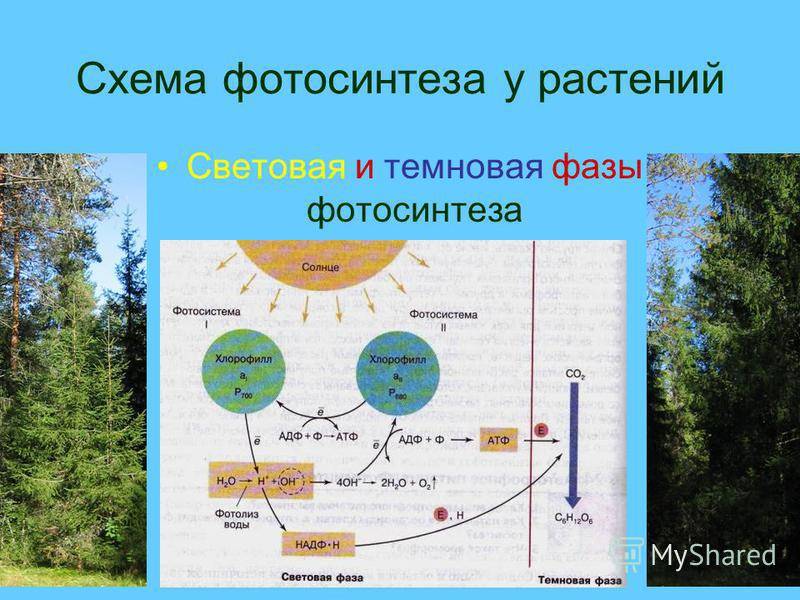 Пигмент ФСII, Р680 – очень сильный окислитель, но его восстановительная сила в возбужденном состоянии Р680* относительно невысока, ее не хватит для восстановления NADPH. Пигмент второй фотосистемы, Р700 – не очень сильный окислитель, но Р700* имеет очень высокую восстановительную способность. Таким образом, роль первой фотосистемы при функционировании полной ЭТЦ – восстановление NADPH. Происходит это после возбуждения P700 вспышкой света («Photon» на рисунке) и ухода электрона по ЭТЦ первой фотосистемы на ферредоксин. Ушедший электрон восполняется электроном, выбитым из второй фотосистемы. По пути переноса электронов от ФСII к ФСI совершается работа – перекачивание протонов, создание градиента, за счет которого далее синтезируется АТФ.
Пигмент ФСII, Р680 – очень сильный окислитель, но его восстановительная сила в возбужденном состоянии Р680* относительно невысока, ее не хватит для восстановления NADPH. Пигмент второй фотосистемы, Р700 – не очень сильный окислитель, но Р700* имеет очень высокую восстановительную способность. Таким образом, роль первой фотосистемы при функционировании полной ЭТЦ – восстановление NADPH. Происходит это после возбуждения P700 вспышкой света («Photon» на рисунке) и ухода электрона по ЭТЦ первой фотосистемы на ферредоксин. Ушедший электрон восполняется электроном, выбитым из второй фотосистемы. По пути переноса электронов от ФСII к ФСI совершается работа – перекачивание протонов, создание градиента, за счет которого далее синтезируется АТФ.
Таким образом, электроны от ФСII восстанавливают Р700+ ФСI, и снова возникает проблема донора электронов, уже для второй фотосистемы. Чисто теоретически донором может быть все тот же сероводород, как у зеленых серных бактерий. Сероводород довольно легко окислить и продукт окисления довольно инертен и неопасен. Но сероводорода мало на Земле. А вот воды много. При этом вода – очень плохой восстановитель, очень трудно отобрать у нее электроны. Более того, продукты окисления высокоактивны и могут вызывать повреждения. Но фактор распространения оказался решающим, вода используется в качестве донора электронов: 2h3O 4e-+4H++O2. Именно очень сильный окислительный потенциал P680 позволяет существовать оксигенному фотосинтезу, который появился где-то 2,1-2,5 млрд лет назад у цианобактерий. Помимо огромного преимущества, для самих фотосинтетиков — это событие сыграло одну из ключевых ролей в формировании атмосферы, что называется кислородной катастрофой, именно оксигеный фотосинтез определил создание условий, пригодных для развития сложных форм жизни на Земле.
Но сероводорода мало на Земле. А вот воды много. При этом вода – очень плохой восстановитель, очень трудно отобрать у нее электроны. Более того, продукты окисления высокоактивны и могут вызывать повреждения. Но фактор распространения оказался решающим, вода используется в качестве донора электронов: 2h3O 4e-+4H++O2. Именно очень сильный окислительный потенциал P680 позволяет существовать оксигенному фотосинтезу, который появился где-то 2,1-2,5 млрд лет назад у цианобактерий. Помимо огромного преимущества, для самих фотосинтетиков — это событие сыграло одну из ключевых ролей в формировании атмосферы, что называется кислородной катастрофой, именно оксигеный фотосинтез определил создание условий, пригодных для развития сложных форм жизни на Земле.
Сам процесс окисления воды происходит в марганцевом кластере – сложноустроенном белковом комплексе, в реакционном центре которого содержится, помимо прочего, 4 марганца, хлор и кальций (см рис 19), связанных с аминокислотными остатками. Кластер может постепенно отдавать по одному электрону на реакционный центр ФСII, восстанавливая пигмент Р680+, отдавший электрон в ЭТЦ.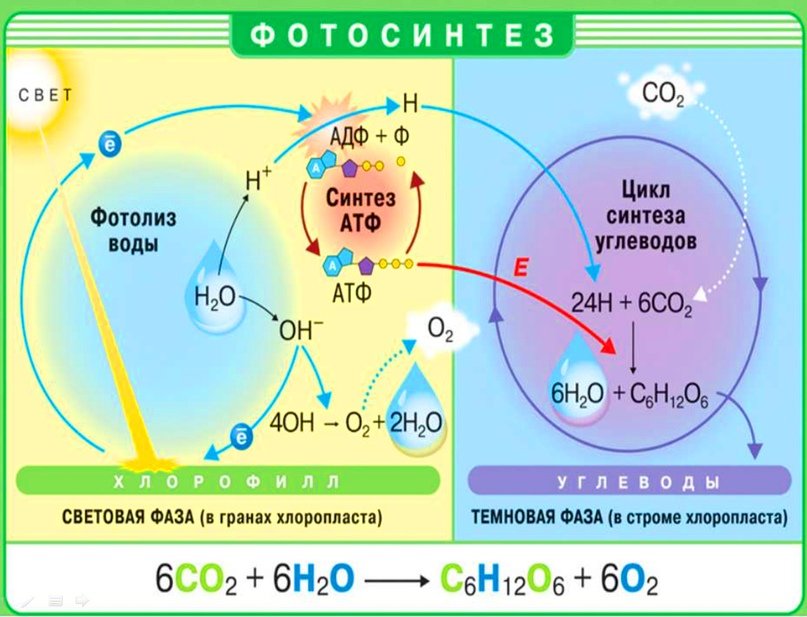 Посредником между Р680 и марганцевым кластером является Tyr Z (тирозин-161 белка D1 второй фотосистемы). Марганцевый кластер, изменяя поочередно степень окисления, может отдавать последовательно 4 электрона, которые далее восполняются за счет окисления двух молекул воды.
Посредником между Р680 и марганцевым кластером является Tyr Z (тирозин-161 белка D1 второй фотосистемы). Марганцевый кластер, изменяя поочередно степень окисления, может отдавать последовательно 4 электрона, которые далее восполняются за счет окисления двух молекул воды.
Марганцевый кластер располагается с люменальной стороны тилакоида, соединен с донорной частью ФСII. Обратите внимание, что в результате работы Mn-кластера в люмене также накапливаются протоны.
Вторая фотосистема (рис 20) состоит из множества белков – более 20, среди них есть белки, в которых непосредственно происходят фотохимические процессы (D1 и D2), комплексы внутренних и внешних антенн и большое количество других, необходимых для сборки, стабилизации, регулирования. Главный белок – D1, на нем происходят практически все основные события – разделение зарядов, формирование донорно-акцепторных пар. Структурно на него очень похож D2, с которым они находятся в димере. Р680 расположен на стыке между двумя этими белками, образуя связь с аминокислотами одного и второго. Всего реакционный центр ФС II несет 2 β-каротина, 2 феофитина, 6 хлорофиллов а.
На D1 и D2 есть похожие цепи электронных переносчиков, но функционирует только та, что на D1. С Р680* электрон передается на первичный акцептор – феофитин, с него на связанный хинон QA, с него на QB, который выходит в мембрану, в пул пластохинонов и далее может идти к цитохромному комплексу.
Забегая вперед, скажем про еще один (альтернативный) путь восстановления P680+. Он реализуется в том случае, если по каким-либо причинам перекрывается перенос электронов по ЭТЦ или нарушается работа водоокисляющего комплекса. В данном случае электрон с феофитина и/или с хинола поступает на цитохром b559, откуда далее идет к хлорофиллу ChlD и/или ChlZ и затем через каротин на P680+.
CP43 (=psbC) и CP47(=psbB) – внутренние светособирающие антенны фотосистемы. В них содержится несколько хлорофиллов а и β-каротины. CP29,26,24 (=Lhcb4,5,6) – внешние антенны, в них содержатся хлорофиллы а и b, лютеины, неоксантины и виолаксантины (которые превращаются в зеаксантин в виолаксантиновом цикле) . Также существует подвижный светособирающий комплекс (ССК II). Состоит из трех субъединиц (Lhcb1,2,3). При тусклой и нормальной освещенности ССК II находится рядом с ФСII, при избыточном освещении ССК II отходит и может присоединяться к ФС I.
Цитохром b6f в общем похож на митохондриальные и бактериальные bc1. Это димерный комплекс, в котором есть два сайта связывания пластохинонов (Qp – со стороноы люмена, здесь окисляется пластохинол; Qn – со стороны стромы, в нем восстанавливается пластохинон). Также имеются цитохромы – низкопотенциальный bL и высокопотенциальный bH, цитохром f – связь с пластоцианином. Подробно прочитать про функционирование можно в Википедии, статья очень хорошо написана (https://ru.wikipedia.org/?oldid=94460479). Если коротко, то основные происходящие в комплексе процессы следующие. Пластохинол связывается в Qp сайте, окисляется, превращаясь в семихинон, при этом первый протон выходит в люмен. Первый электрон идет по так называемой высокопотенциальной ветви на белок Риске – 2Fe2S – содержащий белок, который, претерпевая конформационные изменения после акцептирования электрона, передает его на цитохром f. После передачи электрона на белок Риске этот путь временно перекрывается и следующий электрон уходит от семихинона, окисляя его до пластохинона (второй протон уходит в люмен), идет по цепи цитохромов – низкопотенциального и высокопотенциального, откуда сбрасывается на пластохинон, связанный в Qn сайте, восстанавливая его до семихинона (протон берется из стромы). Из Qp пластохинон уходит обратно в мембрану и в Qp связывается следующий пластохинол. Снова повторяются описанные процессы окисления и переноса электронов, и уже в Qn – сайте семихинон восстанавливается до пластохинола, который может уйти в мембранный пул, а затем связаться в Qp сайте. Описанные процессы называются Q-циклом (рис. 23). Можно представить все в виде уравнения:
2Cytf3++2H++2PQh3+PQ +2e-2PQ+PQh3+4H++2Cytf2+
Сократив хиноны и хинолы, но не сокращая протоны, так как они по разные стороны от мембраны, получим:
2Cytf3++2H++PQh3 +2e-PQ+4H++2Cytf2+
Таким образом, если рассматривать процесс в динамике, получается, что на каждый пластохинол, окисленный в Qp сайте через мембрану переносится 4 протона и восстанавливается 2 цитохрома f. А цитохром f передает электрон на пластоцианин. Значит, на каждый окисленный пластохинол к первой фотосистеме приходит два электрона.
Пластоцианин, приняв электрон от цитохрома f, (Cu2+Pc + e− → Cu+Pc), перемещается в люменальном пространстве и передает e− на I фотосистему: Cu+Pc → Cu2+Pc + e−.
Первая фотосистема также является сложным белковым комплексом. Белки A (PsaA) и В (PsaB) образуют гетеродимер, содержащий реакционный центр и первичные акцепторы: Р700, А0, А1, Fx. Белок С содержит железо-серные кластеры (4Fe4S) FA, FB. Белки D, E образуют сайт связывания ферредоксина (Fd), F обуславливает связь с пластоцианином (PC). С ферредоксином связывается FNR – Fd:NADP+-редуктаза (содержит FAD – кофактор флавиновой природы), восстанавливается NADP+ до NADPH.
В первой фотосистеме, в отличие от второй, функционируют обе ветви транспорта, но, возможно, при разных условиях. Также есть вариант переноса на уровне железосерных кластеров.
Пигмент P700* — очень мощный восстановитель, его потенциала хватает, чтобы восстановить NADP+.
Первичным акцептором электрона является хлорофилл а695 (А0), далее электрон переходит на акцептор хинольной природы (филлохинон, витамин К1) и FeS кластеры.
У первой фотосистемы также есть свои антенны. Внутренние антенны располагаются непосредственно на белках А и B, содержат β-каротины и хлорофилл а. Внешние – Lhca1-4, несут хророфиллы а и b, каротин и ксантофиллы.
Ферредоксин растений содержит 2Fe2S кластер, относительно устойчив к кислороду.
Теперь немного поговорим о том, как может изменяться транспорт электронов, в зависимости от условий. На ЭТЦ влияет множество различных факторов, основные из которых: интенсивность освещения, доступность NADP+, величина протонного градиента, pH стромы и люмена.
На избыточном свету могут начинать происходить фотодеструктивные процессы. Прежде всего это связано с Р680. Если по каким-либо причинам транспорт электронов по ЭТЦ перекрывается, невозможность релаксировать возбужденное состояние P680* приведет к многочисленным, уже вкратце упоминавшимся негативным последствиям, в нейтрализации которых каротиноиды играют существенную роль. Еще вариант восстановления – замена D1 белка, который на ярком свету очень быстро разрушается.
Чтобы снизить риск возникновения фотодеструкции, существует ряд превентивных мер. Мы уже говорили об альтернативном транспорте вокруг ФС II – это одна из таких мер.
Также существует транспорт вокруг первой фотосистемы. ФС I более устойчива на ярком свету, чем ФСII, поэтому при избыточной интенсивности света происходит изменение антенных систем: ССКII отсоединяется от ФСII и присоединяется к ФСI. Это, с одной стороны, приводит к уменьшению притока энергии к Р860, а значит и уменьшает разрушение ФСII. С другой стороны, интенсифицирует приток энергии к Р700. В циклическом транспорте вокруг ФСI участвует новый фермент – ферредоксин-пластохинон оксидоредуктаза, таким образом, электроны с ферредоксина идут не на восстановление NADPH, а на восстановление пластохинонов. NADPH нет, но градиент протонов нагнетается, АТФ синтезируется (работает Q-цикл). Этот транспорт помогает как раз в том случае, когда из-за перевосстановленности пула NADP (нехватки NADP+) происходит торможение потока электронов по ЭТЦ, что может усилить их сброс на кислород с образованием активных форм кислорода. Как уже упоминали, переход ССК II связан с изменением мембранной структуры хлоропласт – разбираются граны. Дело в том, ч в хлоропластах каждый компонент ЭТЦ расположен в определенном месте тилакоидых мембран и мембран ламелл стромы. Из-за сильно выпирающей стромальной части первая фотосистема может располагаться только на ламеллах стромы и на внешних частях гран. ССК II участвуют в сборке гран, взаимодействуя с ССК II других тилакоидов, то есть располагаются на смежных мембранах внутри граны. Там же находится и вторая фотосистема. Именно из-за этого при перераспределении ССК II между тилакоидами граны и ламеллами происходит распад гран. Этот процесс обратим: при изменении интенсивности освещения или других условий граны вновь собираются.
Другой альтернативный вариант, как можно разгрузить ЭТЦ – перенести электроны с ферредоксина на кислород с образованием суперокидрадикала. Это активная форма кислорода, но она может быстро нейтрализоваться до перекиси водорода (другая форма АФК) супероксиддисмутазой. А перекись водорода пероксидазой за счет окисления аскорбата (анион аскорбиновой кислоты) восстанавливается до кислорода и воды. Другое название этого процесса – реакция Мелера.
Существует еще один альтернативный путь – хлородыхание. В тилакоидной мембране присутствует NADPH-дегидрогеназа – фермент, окисляющий NADPH и передающий электроны на пластохиноны. Пластохиноны дальше могут окисляться на пластидной альтернативной оксидазе, в результате работы которой кислород восстанавливается до воды. Таким образом, в ходе фотосинтеза кислород не только выделяется, но и расходуется, при определенных условиях.
Вкратце рассмотрим еще один интересный механизм защиты СФ II от деструкции – виолаксантиновый цикл. Он проходит в антенных комплнксах, содержащих соответствующие ксантофиллы, при избыточной освещенности. Реакции и превращения изображены на рисунке 28. На пути виолаксантин-зеаксантин действует расположенная с люменальной стороны мембраны де-эпоксидаза, в обратном направлении – эпоксидаза, располагающаяся со стромальной стороны мембраны тилакоида. Ферменты регулируются состоянием pH. Де-эпоксидаза активируется при сильном закислении люмена (=стресс, избыточное освещение), то есть при создании довольно высокого протонного градиента, который, в свою очередь, тоже может тормозить ЭТЦ. Конечным результатом реакции является изменение числа сопряженных двойных связей в ксантофилле, что влияет на его фотохимические свойства. На рисунке 29 показаны энергетические уровни (основные и возбужденные) виолаксантина, хлорофилла и зеаксантина, из соотношения которых видно, как может между ними переходить возбужденное состояние. Таким образом, виолаксантин работает как антенна и передает энергию на хлорофилл (при низкой освещенности), а зеаксантин выполняет защитную роль, забирая избыток энергии.
Приложения (Происхождение пластид, строение хлоропласта).
световая фаза, темновая фаза. Хемосинтез»
Тема: «Фотосинтез: световая фаза, темновая фаза. Хемосинтез»
Цели и задачи урока:
Образовательные: сформировать представления об особенностях процессов пластического обмена, световой и темновой фазах фотосинтеза, особенности хемосинтеза.
Воспитательные: формирование научного мировоззрения, осуществление экологического, санитарно — гигиенического воспитания, привитие интереса к естественным наукам.
Развивающие: развивать умения наблюдать, сравнивать, обобщать, делать выводы, предъявлять результаты своей деятельности.
Оборудование: таблица «Фазы фотосинтеза»
Тип урока: изучение нового материала.
Дата:_____________________
Ход урока:
- Организационный момент:
— Приветствие класса.
- Актуализация и мотивация опорных знаний:
— Кто такие автотрофы?
— Что такое фотосинтез?
— Где протекают процессы фотосинтеза?
— Что такое ассимиляция?
III. Изучение нового материала:
Пластический обмен – совокупность процессов синтеза. Основные процессы ассимиляции – синтез белков, жиров, углеводов и нуклеиновых кислот. Так же процессы ассимиляции характерны для фотосинтеза и хемосинтеза
Фотосинтез:
Фотосинтез – образования органических веществ из неорганических благодаря превращению энергии солнца на энергию в химических связях синтезированных углеводов.
6СО2 + 6Н2О + Энергия света = С6Н12О6 + 6О2
Фотосинтез происходит в хлоропластах, благодаря веществу хлорофиллу. Хлорофилл по своей структуре похож на гемоглобин, вместо Ферума у него Магний.
Фазы фотосинтеза:
- Световая фаза – с обязательным участием солнечного света.
- Темновая фаза – солнечный свет необязателен.
Световая фаза:
- Поглощение хлорофиллом кванта (фотона) солнечного света
- Электроны Магния переходят на более высокий энергетический уровень
- Передача энергии другим веществам для синтеза АТФ и восстановления НАДФ
- Распад воды – фотолиз = электроны е — + протоны Н+ + кислород.
- Восстановление НАДФ с помощью протонов Н+
- Синтез АТФ из АДФ.
Световая фаза – совокупность процессов, которые обеспечивают синтез молекулярного кислорода, водорода и АТФ за счет солнечного света.
Темновая фаза:
Реакции протекают круглосуточно в матриксе хлоропластов, без участия солнечного света, в зависимости от потребности в углеводах. Основой темновой фазы является цикл реакций, под названием цикл Кальвина. В темновой фазе из углекислого газа, Водорода и энергии АТФ синтезируется глюкоза.
Темновая фаза – совокупность цикличных реакций, которые благодаря химической энергии АТФ обеспечивают образование глюкозы.
Роль фотосинтеза:
- С автотрофов начинаются все цепи питания;
- Влияние на состав газов атмосферы;
- Оберегает от накапливания СО2 и перегревания планеты.
Задание ст.87, заполнить сравнительную таблицу.
Хемосинтез – процесс синтеза органических веществ из неорганических благодаря энергии, которая освобождается в процессе окисления неорганических веществ. К хемотрофным организмам относятся: нитрифицирующие, метансинтезирующие, железобактерии, и т.д.
— Бактерии – хемосинтетики последовательно окисляют аммиак до солей нитратной кислоты, которые в дальнейшем используются растениями для полноценного питания.
— Железобактерии — окисляют двухвалентное железо до трехвалентного.
— Бесцветные аэробные хемотрофные бактерии – окисляют вещества с Серой до сульфатной кислоты.
Роль хемосинтеза в природе:
- Обеспечение круговорота веществ в природе
- Синтезируют вещества там, куда не попадает солнечный свет.
- Синтезируют природные ресурсы.
IV. Обобщение и систематизация знаний:
— Заполнить таблицу на стр. 91
V. Домашнее задание: §20-21
Фотосинтез значение в природе: световая и темновая стадии синтеза и их продукты
По типу питания живые организмы делятся на автотрофы, гетеротрофы и миксотрофы. Автотрофы (греч. αὐτός — сам + τροφ — пища) — организмы, которые самостоятельно способны синтезировать органические вещества из неорганических. Гетеротрофы (греч. ἕτερος — иной + τροφή — пища) — организмы, использующие для питания готовые органические вещества.
Наконец, миксотрофы (греч. μῖξις — смешение + τροφή — пища) — организмы, которые могут использовать как гетеротрофный, так и автотрофный способ питания. К примеру, эвглена зеленая на свету начинает фотосинтезировать, а в темноте питается гетеротрофно.
Фотосинтез (греч. φῶς — свет и σύνθεσις — синтез) — сложный химический процесс преобразования энергии квантов света в энергию химических связей. В результате фотосинтеза происходит синтез органических веществ из неорганических.
Этот процесс уникален и происходит только в растительных клетках, а также у некоторых бактерий. Фотосинтез осуществляется при участии хлорофилла (греч. χλωρός — зелёный и φύλλον — лист) — зеленого пигмента, окрашивающего органы растений в зеленый цвет. Существуют и другие вспомогательные пигменты, которые вместе с хлорофиллом выполняют светособирающую или светозащитную функции.
Ниже вы увидите сравнение строения хлорофилла и гемоглобина. Обратите внимание, что в центре молекулы хлорофилла находится ион Mg.
В высшей степени гениально значение процесса фотосинтеза подчеркнул русский ученый К.А.
Тимирязев: «Все органические вещества, как бы они ни были разнообразны, где бы они ни встречались, в растении ли, в животном или человеке, прошли через лист, произошли от веществ, выработанных листом.
Вне листа или, вернее, вне хлорофиллового зерна в природе не существует лаборатории, где бы выделялось органическое вещество. Во всех других органах и организмах оно превращается, преобразуется, только здесь оно образуется вновь из вещества неорганического»
Более подробно мы обсудим значение фотосинтеза в завершение этой статьи. Фотосинтез состоит из двух фаз: светозависимой (световой) и светонезависимой (темновой). Я рекомендую использовать названия светозависимая и светонезависимая, так как они способствуют более глубокому (и правильному!) пониманию фотосинтеза.
Светозависимая фаза (световая)
Эта фаза происходит только на свету на мембранах тилакоидов в хлоропластах. В ней принимают участие различные ферменты, белки-переносчики, молекулы АТФ-синтетазы и зеленый пигмент хлорофилл.
Хлорофилл выполняет две функции: поглощения и передачи энергии. При воздействии кванта света хлорофилл теряет электрон, переходя в возбужденное состояние. С помощью переносчиков электроны скапливаются с наружной поверхности мембраны тилакоидов, тем временем внутри тилакоида происходит фотолиз воды (разложение под действием света):
- h3O —> H+ + OH-
- Гидроксид-ионы отдают лишний электрон, превращаясь в реакционно способные радикалы OH, которые собираются вместе и образуют молекулу воды и свободный кислород (это побочный продукт, который в дальнейшем удаляется в ходе газообмена).
- 4OH —> 2h3O + O2↑
Образовавшиеся при фотолизе воды протоны (H+) скапливаются с внутренней стороны мембраны тилакоидов, а электроны — с внешней. В результате по обе стороны мембраны накапливаются противоположные заряды.
При достижении критической разницы, часть протонов проталкивается на внешнюю сторону мембраны через канал АТФ-синтетазы. В результате этого выделяется энергия, которая может быть использована для фосфорилирования молекул АДФ:
Протоны, попав на поверхность мембраны тилакоидов, соединяются с электронами и образуют атомарный водород, который используется для восстановления молекулы-переносчика НАДФ (никотинамиддинуклеотидфосфат). Благодаря этому окисленная форма — НАФД+ превращается в восстановленную — НАДФ∗h3.
Предлагаю создать квинтэссенцию из полученных нами знаний. Итак, в результате светозависимой фазы фотосинтеза образуются:
- Свободный кислород O2 — в результате фотолиза воды
- АТФ — универсальный источник энергии
- НАДФ∗h3 — форма запасания атомов водорода
Кислород удаляется из клетки как побочный продукт фотосинтеза, он совершенно не нужен растению. АТФ и НАДФ∗h3 в дальнейшем оказываются более полезны: они транспортируются в строму хлоропласта и принимают участие в светонезависимой фазе фотосинтеза.
Светонезависимая (темновая) фаза
Светонезависимая фаза происходит в строме (матриксе) хлоропласта постоянно: и днем, и ночью — вне зависимости от освещения.
При участии АТФ и НАДФ∗h3 происходит восстановление CO2 до глюкозы C6h22O6. В светонезависимой фазе происходит цикл Кальвина, в ходе которого и образуется глюкоза. Для образования одной молекулы глюкозы требуется 6 молекул CO2, 12 НАДФ∗h3 и 18 АТФ.
Таким образом, в результате темновой (светонезависимой) фазы фотосинтеза образуется глюкоза, которая в дальнейшем может быть преобразована в крахмал, служащий для запасания питательных веществ у растений.
Значение фотосинтеза
Значение фотосинтеза невозможно переоценить. Уверенно утверждаю: именно благодаря этому процессу жизнь на Земле приобрела такие чудесные и изумительные формы, какие мы видим вокруг себя: удивительные растения, прекрасные цветы и самые разнообразные животные.
В разделе эволюции мы уже обсуждали, что изначально в составе атмосферы Земли не было кислорода: миллиарды лет назад его начали вырабатывать первые фотосинтезирующие бактерии — сине-зеленые водоросли (цианобактерии). Постепенно кислород накапливался, и со временем на Земле стало возможно аэробное (кислородное) дыхание. Возник озоновый слой, защищающий все живое на нашей планете от губительного ультрафиолета.
Говоря о роли фотосинтеза, выделим следующие функции, объединяющиеся в так называемую космическую роль растений. Итак, растения за счет фотосинтеза:
- Синтезируют органические вещества, являющиеся пищей для всего живого на планете
- Преобразуют энергию света в энергию химических связей, создают органическую массу
- Растения поддерживают определенный процент содержания O2 в атмосфере, очищают ее от избытка CO2
- Способствуют образованию защитного озонового экрана, поглощающего губительное для жизни ультрафиолетовое излучение
Хемосинтез (греч. chemeia – химия + synthesis — синтез)
Хемосинтез — автотрофный тип питания, который характерен для некоторых микроорганизмов, способных создавать органические вещества из неорганических. Это осуществляется за счет энергии, получаемой при окислении других неорганических соединений (железо- , азото-, серосодержащих веществ).
Хемосинтез был открыт русским микробиологом С.Н. Виноградским в 1888 году. Большинство хемосинтезирующих бактерий относится к аэробам, для жизни им необходим кислород.
При окислении неорганических веществ выделяется энергия, которую организмы запасают в виде энергии химических связей. Так нитрифицирующие бактерии последовательно окисляют аммиак до нитрита, а затем — нитрата. Нитраты могут быть усвоены растениями и служат удобрением.
Помимо нитрифицирующих бактерий, встречаются:
- Серобактерии — окисляют h3S —> S 0 —> (S+4O3)2- —> (S+6O4)2-
- Железобактерии — окисляют Fe+2 —>Fe+3
- Водородные бактерии — окисляют h3 —> H+12O
- Карбоксидобактерии — окисляют CO до CO2
Значение хемосинтеза
Хемосинтезирующие бактерии являются неотъемлемым звеном круговорота в природе таких элементов как: азот, сера, железо.
Нитрифицирующие бактерии обеспечивают переработку (нейтрализацию) ядовитого вещества — аммиака. Они также обогащают почву нитратами, которые очень важны для нормального роста и развития растений (это происходит за счет клубеньковых бактерий на корнях бобовых растений).
Источник: https://studarium.ru/article/124
Фотосинтез | Биология
Фотосинтез — это преобразование энергии света в энергию химических связей органических соединений.
Фотосинтез характерен для растений, в том числе всех водорослей, ряда прокариот, в том числе цианобактерий, некоторых одноклеточных эукариот.
В большинстве случаев при фотосинтезе в качестве побочного продукта образуется кислород (O2). Однако это не всегда так, поскольку существует несколько разных путей фотосинтеза. В случае выделения кислорода его источником является вода, от которой на нужды фотосинтеза отщепляются атомы водорода.
Фотосинтез состоит из множества реакций, в которых участвуют различные пигменты, ферменты, коферменты и др. Основными пигментами являются хлорофиллы, кроме них — каротиноиды и фикобилины.
В природе распространены два пути фотосинтеза растений: C3 и С4. У других организмов есть своя специфика реакций.
Все, что объединяет эти разные процессы под термином «фотосинтез», – во всех них в общей сложности происходит преобразование энергии фотонов в химическую связь.
Для сравнения: при хемосинтезе происходит преобразование энергии химической связи одних соединений (неорганических) в другие — органические.
Выделяют две фазы фотосинтеза — световую и темновую. Первая зависит от светового излучения (hν), которое необходимо для протекания реакций. Темновая фаза является светонезависимой.
У растений фотосинтез протекает в хлоропластах. В результате всех реакций образуются первичные органические вещества, из которых потом синтезируются углеводы, аминокислоты, жирные кислоты и др. Обычно суммарную реакцию фотосинтеза пишут в отношении глюкозы — наиболее распространенного продукта фотосинтеза:
6CO2 + 6h3O → C6h22O6 + 6O2
Атомы кислорода, входящие в молекулу O2, берутся не из углекислого газа, а из воды. Углекислый газ – источник углерода, что более важно. Благодаря его связыванию у растений появляется возможность синтеза органики.
Представленная выше химическая реакция есть обобщенная и суммарная. Она далека от сути процесса. Так глюкоза не образуется из шести отдельных молекул углекислоты. Связывание CO2 происходит по одной молекуле, которая сначала присоединяется к уже существующему пятиуглеродному сахару.
Для прокариот характерны свои особенности фотосинтеза. Так у бактерий главный пигмент — бактериохлорофилл, и не выделяется кислород, так как водород берется не из воды, а часто из сероводорода или других веществ. У сине-зеленых водорослей основным пигментом является хлорофилл, и при фотосинтезе выделяется кислород.
Световая фаза фотосинтеза
В световой фазе фотосинтеза происходит синтез АТФ и НАДФ·h3 за счет лучистой энергии. Это происходит на тилакоидах хлоропластов, где пигменты и ферменты образуют сложные комплексы для функционирования электрохимических цепей, по которым передаются электроны и отчасти протоны водорода.
Электроны в конечном итоге оказываются у кофермента НАДФ, который, заряжаясь отрицательно, притягивает к себе часть протонов и превращается в НАДФ·h3. Также накопление протонов по одну сторону тилакоидной мембраны и электронов по другую создает электрохимический градиент, потенциал которого используется ферментом АТФ-синтетазой для синтеза АТФ из АДФ и фосфорной кислоты.
Главными пигментами фотосинтеза являются различные хлорофиллы. Их молекулы улавливают излучение определенных, отчасти разных спектров света. При этом некоторые электроны молекул хлорофилла переходят на более высокий энергетический уровень.
Это неустойчивое состояние, и по-идее электроны путем того же излучения должны отдать в пространство полученную из вне энергию и вернуться на прежний уровень.
Однако в фотосинтезирующих клетках возбужденные электроны захватываются акцепторами и с постепенным уменьшением своей энергии передаются по цепи переносчиков.
На мембранах тилакоидов существуют два типа фотосистем, испускающих электроны при действия света. Фотосистемы представляют собой сложный комплекс большей частью хлорофильных пигментов с реакционным центром, от которого и отрываются электроны. В фотосистеме солнечный свет ловит множество молекул, но вся энергия собирается в реакционном центре.
Электроны фотосистемы I, пройдя по цепи переносчиков, восстанавливают НАДФ.
Энергия электронов, оторвавшихся от фотосистемы II, используется для синтеза АТФ. А сами электроны фотосистемы II заполняют электронные дырки фотосистемы I.
Дырки второй фотосистемы заполняются электронами, образующимися в результате фотолиза воды.
Фотолиз также происходит при участии света и заключается в разложении h3O на протоны, электроны и кислород. Именно в результате фотолиза воды образуется свободный кислород.
Протоны участвуют в создании электрохимического градиента и восстановлении НАДФ. Электроны получает хлорофилл фотосистемы II.
Примерное суммарное уравнение световой фазы фотосинтеза:
h3O + НАДФ + 2АДФ + 2Ф → ½O2 + НАДФ · h3 + 2АТФ
Циклический транспорт электронов
Выше описана так называемый нецикличная световая фаза фотосинтеза. Есть еще циклический транспорт электронов, когда восстановления НАДФ не происходит.
При этом электроны от фотосистемы I уходят на цепь переносчиков, где идет синтез АТФ. То есть эта электрон-транспортная цепь получает электроны из фотосистемы I, а не II.
Первая фотосистема как бы реализует цикл: в нее возвращаются ей же испускаемые электроны. По дороге они тратят часть своей энергии на синтез АТФ.
Фотофосфорилирование и окислительное фосфорилирование
Световую фазу фотосинтеза можно сравнить с этапом клеточного дыхания — окислительным фосфорилированием, которое протекает на кристах митохондрий. Там тоже происходит синтез АТФ за счет передачи электронов и протонов по цепи переносчиков.
Однако в случае фотосинтеза энергия запасается в АТФ не для нужд клетки, а в основном для потребностей темновой фазы фотосинтеза. И если при дыхании первоначальным источником энергии служат органические вещества, то при фотосинтезе – солнечный свет.
Синтез АТФ при фотосинтезе называется фотофосфорилированием, а не окислительным фосфорилированием.
Темновая фаза фотосинтеза
Впервые темновую фазу фотосинтеза подробно изучили Кальвин, Бенсон, Бэссем. Открытый ими цикл реакций в последствии был назван циклом Кальвина, или C3-фотосинтезом. У определенных групп растений наблюдается видоизмененный путь фотосинтеза – C4, также называемый циклом Хэтча-Слэка.
В темновых реакциях фотосинтеза происходит фиксация CO2. Темновая фаза протекает в строме хлоропласта.
Восстановление CO2 происходит за счет энергии АТФ и восстановительной силы НАДФ·h3, образующихся в световых реакциях. Без них фиксации углерода не происходит. Поэтому хотя темновая фаза напрямую не зависит от света, но обычно также протекает на свету.
Цикл Кальвина
Первая реакция темновой фазы – присоединение CO2 (карбоксилирование) к 1,5-рибулезобифосфату (рибулезо-1,5-дифосфат) – РиБФ. Последний представляет собой дважды фосфорилированную рибозу. Данную реакцию катализирует фермент рибулезо-1,5-дифосфаткарбоксилаза, также называемый рубиско.
В результате карбоксилирования образуется неустойчивое шестиуглеродное соединение, которое в результате гидролиза распадается на две трехуглеродные молекулы фосфоглицериновой кислоты (ФГК) – первый продукт фотосинтеза. ФГК также называют фосфоглицератом.
РиБФ + CO2 + h3O → 2ФГК
ФГК содержит три атома углерода, один из которых входит в состав кислотной карбоксильной группы (-COOH):
Из ФГК образуется трехуглеродный сахар (глицеральдегидфосфат) триозофосфат (ТФ), включающий уже альдегидную группу (-CHO):
ФГК (3-кислота) → ТФ (3-сахар)
На данную реакцию затрачивается энергия АТФ и восстановительная сила НАДФ · h3. ТФ — первый углевод фотосинтеза.
После этого большая часть триозофосфата затрачивается на регенерацию рибулозобифосфата (РиБФ), который снова используется для связывания CO2. Регенерация включает в себя ряд идущих с затратой АТФ реакций, в которых участвуют сахарофосфаты с количеством атомов углерода от 3 до 7.
В таком круговороте РиБФ и заключается цикл Кальвина.
Из цикла Кальвина выходит меньшая часть образовавшегося в нем ТФ. В перерасчете на 6 связанных молекул углекислого газа выход составляет 2 молекулы триозофосфата. Суммарная реакция цикла с входными и выходными продуктами:
6CO2 + 6h3O → 2ТФ
При этом в связывании участвую 6 молекул РиБФ и образуется 12 молекул ФГК, которые превращаются в 12 ТФ, из которых 10 молекул остаются в цикле и преобразуются в 6 молекул РиБФ.
Поскольку ТФ — это трехуглеродный сахар, а РиБФ — пятиуглеродный, то в отношении атомов углерода имеем: 10 * 3 = 6 * 5. Количество атомов углерода, обеспечивающих цикл не изменяется, весь необходимый РиБФ регенерируется.
А шесть вошедших в цикл молекул углекислоты затрачиваются на образование двух выходящих из цикла молекул триозофосфата.
На цикл Кальвина в расчете на 6 связанных молекул CO2 затрачивается 18 молекул АТФ и 12 молекул НАДФ · h3, которые были синтезированы в реакциях световой фазы фотосинтеза.
Расчет ведется на две выходящие из цикла молекулы триозофосфата, так как образующаяся в последствии молекула глюкозы, включает 6 атомов углерода.
Триозофосфат (ТФ) — конечный продукт цикла Кальвина, но его сложно назвать конечным продуктом фотосинтеза, так как он почти не накапливается, а, вступая в реакции с другими веществами, превращается в глюкозу, сахарозу, крахмал, жиры, жирные кислоты, аминокислоты. Кроме ТФ важную роль играет ФГК. Однако подобные реакции происходят не только у фотосинтезирующих организмов. В этом смысле темновая фаза фотосинтеза – это то же самое, что цикл Кальвина.
Из ФГК путем ступенчатого ферментативного катализа образуется шестиуглеродный сахар фруктозо-6-фосфат, который превращается в глюкозу. В растениях глюкоза может полимеризоваться в крахмал и целлюлозу. Синтез углеводов похож на процесс обратный гликолизу.
Фотодыхание
Кислород подавляет фотосинтез. Чем больше O2 в окружающей среде, тем менее эффективен процесс связывания CO2. Дело в том, что фермент рибулозобифосфат-карбоксилаза (рубиско) может реагировать не только с углекислым газом, но и кислородом. В этом случае темновые реакции несколько иные.
Содержащая пять атомов углерода молекула рибулозобифосфата реагирует уже не с CO2, а с O2. В результате чего образуются по одной молекуле фосфогликолата (C2) и фосфоглицериновой кислоты (C3), а не две ФГК как обычно.
Фосфогликолат — это фосфогликолевая кислота. От нее сразу отщепляется фосфатная группа, и она превращается в гликолевую кислоту (гликолат). Для его «утилизации» снова нужен кислород. Поэтому чем больше в атмосфере кислорода, тем больше он будет стимулировать фотодыхание и тем больше растению будет требоваться кислорода, чтобы избавиться от продуктов реакции.
Фотодыхание — это зависимое от света потребление кислорода и выделение углекислого газа. То есть обмен газов происходит как при дыхании, но протекает в хлоропластах и зависит от светового излучения. От света фотодыхание зависит лишь потому, что рибулозобифосфат образуется только при фотосинтезе.
- При фотодыхании происходит возврат атомов углерода из гликолата в цикл Кальвина в виде фосфоглицериновой кислоты (фосфоглицерата).
- 2 Гликолат (С2) → 2 Глиоксилат (С2) →2 Глицин (C2) — CO2 → Серин (C3) →Гидроксипируват (C3) → Глицерат (C3) → ФГК (C3)
- Как видно, возврат происходит не полный, так как один атом углерода теряется при превращении двух молекул глицина в одну молекулу аминокислоты серина, при этом выделяется углекислый газ.
- Кислород необходим на стадиях превращения гликолата в глиоксилат и глицина в серин.
Превращения гликолата в глиоксилат, а затем в глицин происходят в пероксисомах, синтез серина в митохондриях. Серин снова поступает в пероксисомы, где из него сначала получается гидрооксипируват, а затем глицерат. Глицерат уже поступает в хлоропласты, где из него синтезируется ФГК.
Фотодыхание характерно в основном для растений с C3-типом фотосинтеза. Его можно считать вредным, так как энергия бесполезно тратится на превращения гликолата в ФГК.
Видимо фотодыхание возникло из-за того, что древние растения были не готовы к большому количеству кислорода в атмосфере.
Изначально их эволюция шла в атмосфере богатой углекислым газом, и именно он в основном захватывал реакционный центр фермента рубиско.
C4-фотосинтез, или цикл Хэтча-Слэка
- Если при C3-фотосинтезе первым продуктом темновой фазы является фосфоглицериновая кислота, включающая три атома углерода, то при C4-пути первыми продуктами являются кислоты, содержащие четыре атома углерода: яблочная, щавелевоуксусная, аспарагиновая.
- С4-фотосинтез наблюдается у многих тропических растений, например, сахарного тростника, кукурузы.
- С4-растения эффективнее поглощают оксид углерода, у них почти не выражено фотодыхание.
Растения, в которых темновая фаза фотосинтеза протекает по C4-пути, имеют особое строение листа. В нем проводящие пучки окружены двойным слоем клеток. Внутренний слой — обкладка проводящего пучка. Наружный слой — клетки мезофилла.
Хлоропласты клеток слоев отличаются друг от друга.
Для мезофильных хлоропласт характерны крупные граны, высокая активность фотосистем, отсутствие фермента РиБФ-карбоксилазы (рубиско) и крахмала. То есть хлоропласты этих клеток адаптированы преимущественно для световой фазы фотосинтеза.
В хлоропластах клеток проводящего пучка граны почти не развиты, зато высока концентрация РиБФ-карбоксилазы. Эти хлоропласты адаптированы для темновой фазы фотосинтеза.
Углекислый газ сначала попадает в клетки мезофилла, связывается с органическими кислотами, в таком виде транспортируется в клетки обкладки, освобождается и далее связывается также, как у C3-растений. То есть C4-путь дополняет, а не заменяет C3.
В мезофилле CO2 присоединяется к фосфоенолпирувату (ФЕП) с образованием оксалоацетата (кислота), включающего четыре атома углерода:
Реакция происходит при участии фермента ФЕП-карбоксилазы, обладающего более высоким сродством к CO2, чем рубиско. К тому же ФЕП-карбоксилаза не взаимодействует с кислородом, а значит не затрачивается на фотодыхание.
Таким образом, преимущество C4-фотосинтеза заключается в более эффективной фиксации углекислоты, увеличению ее концентрации в клетках обкладки и следовательно более эффективной работе РиБФ-карбоксилазы, которая почти не расходуется на фотодыхание.
Оксалоацетат превращается в 4-х углеродную дикарбоновую кислоту (малат или аспартат), которая транспортируется в хлоропласты клеток обкладки проводящих пучков. Здесь кислота декарбоксилируется (отнятие CO2), окисляется (отнятие водорода) и превращается в пируват. Водород восстанавливает НАДФ. Пируват возвращается в мезофилл, где из него регенерируется ФЕП с затратой АТФ.
Оторванный CO2 в хлоропластах клеток обкладки уходит на обычный C3-путь темновой фазы фотосинтеза, т. е. в цикл Кальвина.
Фотосинтез по пути Хэтча-Слэка требует больше энергозатрат.
Считается, что C4-путь возник в эволюции позже C3 и во многом является приспособлением против фотодыхания.
Источник: https://biology.su/molecular/photosynthesis
§ 27. Фотосинтез
Понятие фотосинтеза. Для живых организмов Земли основным источником энергии является солнечный свет, благодаря которому прямо или косвенно удовлетворяются их энергетические потребности.
В процессе фотосинтеза растения, водоросли, цианобактерии с помощью специальных пигментов поглощают солнечную энергию и преобразуют ее в энергию химических связей органических веществ. При этом исходными соединениями для синтеза органических веществ служат бедные энергией неорганические вещества — углекислый газ и вода.
Таким образом, фотосинтез (от греч. фотос — свет) — это процесс образования органических веществ из углекислого газа и воды на свету при участии фотосинтетических пигментов.
Фотосинтетические пигменты — это органические вещества, способные поглощать энергию света. При этом они поглощают свет определенной длины волны, а другие световые волны отражают. Различают три класса фотосинтетических пигментов — хлорофиллы, каротиноиды и фикобилины.
Наиболее важными пигментами являются хлорофиллы (от греч. хлорос — зеленый, фил-лон — лист). Известно несколько форм хлорофилла (а, 6, с и др.), различающихся строением молекул. Все хлорофиллы интенсивно поглощают красный и синий свет, а зеленый отражают, что и определяет зеленую окраску этих пигментов, а следовательно, и листьев растений.
Другую группу фотосинтетических пигментов составляют ка р оти н о и ды (от лат. карота — морковь), имеющие различную окраску — оранжевую, желтую, красную и др. Они содержатся в хлоропластах и хромопластах.
Осенью, когда хлорофиллы разрушаются, каротиноиды становятся хорошо заметными (листья меняют окраску). Каротиноиды поглощают свет, недоступный другим пигментам, и передают поглощенную энергию хлорофиллам.
Для красных водорослей и цианобактерий, кроме хлорофиллов, характерны также фикобилины (от греч. фи кос — водоросль и лат. билис — желчь). Эти пигменты имеют красную или синюю окраску и способны поглощать зеленый, синий и фиолетовый свет, проникающий в глубину морей и океанов.
Образованные в ходе фотосинтеза органические соединения являются не только источником энергии, но и источником атомов углерода, водорода и кислорода для синтеза веществ, необходимых организму при построении новых клеток и структур. Значительная часть продуктов фотосинтеза может преобразовываться и запасаться в виде крахмала, жиров или белков.
У растений и водорослей фотосинтез осуществляется в специальных органоидах — хлоропластах. Вы уже знаете, что внутренняя мембрана хлоропластов образует тилакоиды — плоские мешочки, уложенные в стопки (граны). В мембранах тилакоидов расположены особые пигмент-белковые комплексы — фотосистемы.
Существует два типа фотосистем — фотосистема I и фотосистема II. В состав каждой входит светособирающая антенна, образованная молекулами пигментов, реакционный центр и переносчики электронов.
Светособирающая антенна функционирует наподобие воронки: молекулы пигментов поглощают свет и передают всю собранную энергию в реакционный центр, где находится молекула-ловушка, представленная хлорофиллом а (рис. 63).
Поглотив энергию, молекула-ловушка переходит в возбужденное состояние и отдает один из своих электронов специальному переносчику, т. е. окисляется.
Главное различие фотосистем заключается в том, что в состав фотосистемы II входит особый ферментный комплекс, осуществляющий на свету фотолиз воды —расщепление молекул воды с образованием кислорода (02), электронов и протонов (Н+):
2Н20 ->• 02 + 4ё + 4Н+.
Полученные при этом электроны используются фотосистемой II для восстановления молекулы-ловушки в реакционном центре. В фотосистеме I отсутствует подобный ферментный комплекс, и, следовательно, она не способна использовать воду в качестве источника электронов для восстановления своей молекулы-ловушки.
Фотосинтез происходит в две фазы — световую и темновую (рис. 64). Световая фаза осуществляется на мембранах тилакоидов и только при наличии света. Реакции темновой фазы протекают в строме хлоропласта и не требуют с в ета, однако для их прохождения необходимы продукты световой фазы. Поэтому темновая фаза идет практически одновременно со световой.
Световая фаза фотосинтеза. Процессы, протекающие в световой фазе, можно представить следующим образом.
1. Пигменты обеих фотосистем поглощают свет, полученная энергия передается в реакционные центры на молекулы хлорофилла а (молекулы-ловушки), которые переходят в возбужденное состояние и отдают электроны переносчикам.
Электрон из фотосистемы I транспортируется переносчиками на внешнюю сторону тилакоида. Электрон из фотосистемы II с помощью переносчиков доставляется в фотосистему I и восстанавливает молекулу-ловушку в реакционном центре. Так фотосистема I восстанавливается за счет электронов из фотосистемы II, которая, в свою очередь, получает электроны, как вы уже знаете, за счет фотолиза воды.
Кислород, который образуется при фотолизе воды, выделяется из хлоропласта в гиалоплазму клетки, затем в окружающую среду, а протоны (Н+) накапливаются внутри тилакоида.
2. Накопление протонов внутри тилакоида ведет к возникновению электрохимического потенциала на его мембране. В мембране тилакоида содержится фермент АТФ-синтетаза. Когда концентрация протонов достигает определенного уровня, они устремляются в строму хлоропласта, проходя через специальные каналы АТФ-синтетазы. При этом АТФ-синтетаза использует энергию движения протонов для синтеза АТ Ф.
3. На внешней стороне тилакоида происходит восстановление НАДФ+ за счет присоединения к нему электронов и протонов. НАДФ — никотинамид-адениндинуклеотидфосфат (полное название приводится не для запоминания) — переносчик атомов водорода в процессе фотосинтеза.
НАДФ+ + 2ё + 2Н+ ->• НАДФ-Н+Н+.
Таким образом, в ходе световой фазы энергия света поглощается и преобразуется в энергию макроэргических связей АТФ, происходит расщепление воды с выделением кислорода и накопление атомов водорода (в форме НАДФ’Н+Н+). Продуктами световой фазы фотосинтеза являются АТФ, восстановленный НАДФ и кислород. Кислород — побочный продукт фотосинтеза, он выделяется в окружающую среду. АТФ и НАДФ*Н+Н+ используются в темновой фазе фотосинтеза.
Темповая фаза фотосинтеза. Из окружающей среды в хлоропласта поступает углекислый газ, а в строме хлоропластов происходит его восстановление до органических веществ. Это сложный многоступенчатый процесс, который можно выразить общим уравнением:
6С02 + 12НАДФ-Н+Н+ + 18АТФ ->• С6Н1206 + 12НАДФ+ + 18АДФ + 18Н3Р04.
Из приведенного уравнения видно, что для синтеза одной молекулы глюкозы необходимо окислить 12 молекул НДДФ*Н+Н+ (служит источником атомов водорода) и расщепить 18 молекул АТФ (служит источником энергии для синтеза глюкозы). Таким образом, в темновой фазе фотосинтеза энергия макроэргиче-ских связей АТФ преобразуется в энергию химических связей органических веществ.
В темновой фазе фотосинтеза, как уже отмечалось ранее, используются продукты световой фазы (НАДФ’Н+Н+и АТФ), поэтому реакции темновой фазы проходят почти одновременно с реакциями световой фазы. Если объединить процессы, протекающие в обеих фазах, исключив все промежуточные стадии и вещества, можно получить суммарное уравнение процесса фотосинтеза:
- свет
- 6С02 + 6Н20-> С6Н1206 + 602.
- пигменты
Значение фотосинтеза. Уникальность и биологическое значение фотосинтеза определяются тем, что жизнь на нашей планете всем своим существованием обязана этому процессу.
Фотосинтез является основным источником питательных веществ для живых организмов, а также единственным поставщиком свободного кислорода на Земле.
Из кислорода сформировался и поддерживается озоновый слой, защищающий живые организмы Земли от губительного воздействия коротковолнового ультрафиолетового излучения. Кроме того, благодаря фотосинтезу поддерживается относительно постоянное содержание С02 в атмосфере.
В изучение процесса фотосинтеза, раскрытие его механизма большой вклад внесли ученые разных стран: выдающийся русский ученый К- А. Тимирязев, американец М. Кальвин, австралийцы М. Д. Хетч и Р. Ч. Слэк, а также белорусские ученые Т. Н. Годнев и А. А. Шлык.
1. Фотосинтез относится к процессам пластического или энергетического обмена? Почему?
2. В каких органоидах растительной клетки происходит фотосинтез? Что представляет собой фотосистема? Какую функцию выполняют фотосистемы?
3. Каково значение фотосинтеза на Земле? Почему без фототрофных организмов существование биосферы было бы невозможным?
4. Охарактеризуйте световую и темновую фазы фотосинтеза по плану: 1) место протекания; 2) исходные вещества; 3) происходящие процессы; 4) конечные продукты. Какие продукты световой фазы фотосинтеза используются в темновой фазе?
5. Сравните фотосинтез и аэробное дыхание. Укажите черты сходства и различия.
6. Человек за сутки потребляет примерно 430 г кислорода. Дерево средней величины поглощает около 30 кг углекислого газа в год. Сколько деревьев необходимо, чтобы обеспечить одного человека кислородом?
7. Исследователи разделили растения пшеницы на две группы и выращивали их в лаборатории в одинаковых условиях, за исключением того, что растения первой группы освещали красным светом, а растения второй группы — зеленым. У растений какой группы фотосинтез протекал более интенсивно? С чем это связано?
8. С помощью какого эксперимента можно доказать, что кислород, выделяющийся при фотосинтезе, образуется именно из молекул воды, а не из молекул углекислого газа или какого-либо другого вещества?
Биология: учеб. для 10-го кл. учреждений общ. сред, образования с рус. яз. обуч. / Н. Д. Лисов [и др.]; под ред. Н. Д. Лисова. — 3-е изд., перераб. — Минск : Народная асвета, 2014. — 270 с.: ил.
Источник: https://botana.biz/uchebnik/biologiya/10/by001/p027.html
Тест ЕГЭ Биология 11 класс Получить доступ за 75 баллов Фазы фотосинтеза. Световые и темновые реакции фотосинтеза, их взаимосвязь
Фототрофы- организмы, которые обеспечивают себя органическими веществами за счет процесса фотосинтеза. Соответственно, если они обеспечивают сами себя, то вправе называться автотрофами, точнее фотоавтотрофами.
- Фотоавтотрофами являются большинство растений и некоторые бактерии.
- Фотосинтез- процесс синтеза органических веществ из неорганических (воды и углекислого газа) с использованием энергии солнечного света.
- Фотосинтез- это пример пластического обмена, так как идет процесс образования органических веществ.
- Фотосинтез выражается следующим сумарным уранением:
- 6 СО2 + 6 Н2О = С6Н12О6 + 6 О2
- Из уравнения фотосинтеза видно, что из простых веществ углекислого газа и воды в клетках растений образуется глюкоза.
- Помимо глюкозы образуется и кислород, но он является побочным продуктом световой фазы фотосинтеза, зато необходим всем живым существам на Земле.
Результаты фотосинтеза вы можете прочувствовать на себе, когда употребляете в пищу фрукты и ощущаете их сладкий вкус. Этот вкус есть результат фотосинтеза в листьях растений, при котором образовавшаяся фруктоза или глюкоза постепенно накопилась в плодах.
Крахмал в клубнях картофеля тоже является результатом процесса фотосинтеза.
Поэтому чем крупнее листья картофеля, тем больше света они улавливают, следовательно больше образуется глюкозы, которая быстро преобразуется в крахмал.
При выращивании любого растения необходимо ухаживать за ним: соблюдать требования к освещению, удалять сорники, вовремя осуществлять полив и другие меры по уходу. Все это для того, чтобы процесс фотосинтеза шел интенсивно и растение могло активно накапливать органические вещества.
Вы можете услышать такое выражение, как «космическая роль растений».
Первым употребил это понятие русский ученый К.А.Тимирязев (1843-1920).
Он писал: «Это процесс, от которого в конечной инстанции зависят все проявления жизни на нашей планете, а, следовательно, и благосостояние всего человечества».
- Только растения научились воспринимать солнечную энергию, преобразовывать ее в энергию химических связей органических молекул.
- Таким образом, растения дают пищу почти всему остальному живому миру на Земле, а также кислород, который является обязательным участником окислительных реакций, необходимых для образования энергии в клетках живых существ.
- Конечно, исходя из уравнения мы не можем понять механизма процесса фотосинтеза, поэтому в этом разделе кратко попытаемся объяснить этот сложный процесс, происходящий в каждой клетке зеленого растения.
- Если обратится к теме строение растительной клетки, мы вспомним, что в клетках растений есть такой органоид, как хлоропласт.
- В хлоропластах, а конкретно в тилакоидах, находится пигмент хлорофилл, именно ему принадлежит основная роль в фотосинтезе.
- Хлорофилл обладает особой химической структурой, которая позволяет ему улавливать кванты света и переходить в возбужденное состояние.
- Процесс фотосинтеза включает две последовательные и взаимозависящие друг от друга фазы:
- световая фаза фотосинтеза
- темновая фаза фотосинтеза
- Световая фаза идет в гранах (тилакоидах) хлоропластов при обязательном участии солнечного света.
- Молекула восстановленного хлорофилла способна возбуждаться под действием солнечного света и отдавать свои электроны, которые необходимы для создания молекул АТФ.
- Запомните эту схему:
- Возмещение недостающих электронов в хлорофилле происходит благодаря распаду молекул воды.
- Распад молекул воды называется фотолиз.
- При фотолизе также происходит образование кислорода, который как побочный продукт реакции распада воды выходит в окружающую среду.
- Атомный водород, который освобождается также при фотолизе воды, подхватывает переносчик водорода НАДФ+ и востанавливается до НАДФ·2Н.
- В результате перемещений электронов и создания на мембране тилакоида градиента концентрации протонов происходит образование молекул АТФ из АДФ.
- Суммарно это выглядит так:
- Н2О + НАДФ++ 2 АДФ + 2 Фн = НАДФ·2Н + 2 АТФ + ½ О2↑
- Условие протекания данной реакции- воздействие энергии света на хлорофилл.
- Как мы видим, в результате световой фазы фотосинтеза происходят три основных процесса:
- синтез АТФ
- создание НАДФ·2Н
- образование свободного кислорода
- То есть в реакциях световой фазы фотосинтеза накапливается энергия в виде НАДФ·2Н и АТФ, которая тратится в процессах темновой фазы.
- Темновая фаза происходит в строме хлоропласта и протекает независимо от света, но с обязательным участием углекислого газа.
- Углекислый газ попадает в растение через устьица.
- Вспомним строение устьиц:
- Далее углекислый газ претепевает ряд последовательных превращений, чтобы превратиться в глюкозу.
- Для этих реакций требуется большое количество энергии, которая, как мы уже знаем, запаслась в световую фазу фотосинтеза.
- Превращение углекислого газа в глюкозу в ходе темновой фазы фотосинтеза получило название цикла Кальвина.
- Следует отметить, что кроме молекул глюкозы в строме хлоропластов происходит образование и других органических веществ, к примеру, аминокислот, спиртов.
- Результатом темновых реакций является превращение углекислого газа в глюкозу за счет энергии АТФ и НАДФ·2Н; далее глюкоза может запасаться в виде крахмала.
- Опыт, доказывающий, что растениям необходим солнечный свет для фотосинтеза и результатом фотосинтеза являются углеводы:
Существуют и такие растения, которые далеко не сладкие, хотя в них также просходит процесс фотосинтеза. Почему одни сладкие, а в других нет сладкого вкуса?
Дело в том, что в листьях таких растений так же вырабатывается сахар, но из него в процессе метаболизма может образоваться крахмал и большое число других органических соединений, которые служат исходными субстратами для синтеза липидов, аминокислот, нуклеотидов — все они не имеют сладкого привкуса.
Обобщающая таблица фазы фотосинтеза:
| Признаки сравнения | Световая фаза | Темная фаза |
| Условия протекания реакций | Обязательно наличие солнечного света | Солнечный свет не обязателен, может идти днем и ночью |
| Место протекания реакций в хлорпластах | На мембранах тилакоидов (на граннах) | В строме хлоропластов |
| Источник энергии | Солнечный свет | Энергия АТФ, НАДФ·2Н |
| Исходные вещества | Вода, АДФ, Фн, НАДФ+ | Углекислый газ, АТФ, НАДФ·2Н |
| Конечные продукты реакции | Кислород, АТФ, НАДФ·2Н | Глюкоза, АДФ, НАДФ+ |
Источник: https://ladle.ru/education/biologiya/ege/fazy-fotosinteza-ege
Фотосинтез
Фотосинтез — Образование высшими растениями сложных органических веществ из простых соединений — углекислого газа и воды — за счет световой энергии, поглощаемой хлорофиллом. Создаваемые в процессе фотосинтеза органические вещества необходимы растениям для построения их органов и поддержания жизнедеятельности.
Исходные вещества для фотосинтеза — углекислый газ, поступающий в листья из воздуха, и вода — представляют собой продукты полного окисления углерода (CO2) и водорода (h3O).
В образуемых при фотосинтезе органических веществах углерод находится в восстановленном состоянии.
При фотосинтезе система СO2 — Н2O, состоящая из окисленных веществ и находящаяся на низком энергетическом уровне, восстанавливается в менее устойчивую систему СН2O — O2, находящуюся на более высоком энергетическом уровне.
Из уравнения видно, что на получение одной грамм — молекулы глюкозы (С6НО6) расходуется световая энергия в количестве 2872,14 кДж, которая запасается в виде химической энергии. При этом в атмосферу выделяется свободный кислород.
Приведенное уравнение дает конкретное представление о начальных и конечных веществах, участвующих в фотосинтезе, но оно не вскрывает сущности очень сложного биохимического процесса. История учения об углеродном питании растений насчитывает более 200 лет. В трактате «Слово о явлениях воздушных» М. В. Ломоносов в 1753 г. писал, что растение строит свое тело из окружающего его воздуха, поглощенного при помощи листьев. Однако открытие фотосинтеза связывают с именем английского химика Дж. Пристли, который в 1771 г. обнаружил, что на свету зеленые растения «исправляют» воздух, «испорченный» горением. Последующими работами голландского ученого Я. Ингенхауза (1779, 1798 гг.), швейцарских Ж. Сенебье (1782, 1783 гг.) и Н. Соссюра (1804 г.) было установлено, что на свету зеленые растения усваивают из окружающей атмосферы углекислый газ и выделяют кислород. Важную роль в изучении фотосинтеза имели работы К. А. Тимирязева, который показал, что свет является источником энергии для синтеза органических веществ из углекислого газа и воды, и установил максимум поглощения хлорофилла в красной и сине — фиолетовой областях спектра. Дальнейшие исследования многих ученых с использованием современных методов позволили вскрыть многие звенья сложной цепи превращений веществ в растительном организме. Было установлено, что фотосинтез протекает в двух фазах. Первая из них — световая, вторая — темновая. Первая фаза идет только на свету, тогда как вторая — с равным успехом как в темноте, так и на свету. Световая фаза протекает в зеленой фракции хлоропласта — гранах, а все превращения темновой фазы проходят в его бесцветной фракции — цитоплазматическом матриксе. Световая фаза характерна только для фотосинтезирующих клеток, тогда как большинство реакций, составляющих процесс фиксации углекислоты в темновой фазе, свойствен не только фотосинтезирующим клеткам. Световая фаза фотосинтеза начинается с поглощения света пигментами. В химических реакциях световой фазы участвуют лишь молекулы хлорофилла а, находящиеся в активированном (за счет поглощения световой энергии) состоянии. Остальные пигменты — хлорофилл b и каротиноиды — улавливают свет с помощью особых систем, передают полученную энергию на молекулы хлорофилла а. Важнейшая роль световой фазы состоит в построении молекулы АТФ (аденозинтрифосфата), в которой запасается энергия. Процесс образования АТФ в хлоропластах с затратой солнечной энергии называется циклическим фосфорилированием. При распаде АТФ до АДФ (аденозиндифосфата) выделяется около 40 кДж энергии. Для восстановления молекулы НАДФ (никотинамидадениндинуклеотидфосфат) требуется два атома водорода, который получается из воды с помощью света. Активированный светом хлорофилл тратит свою энергию на разложение воды, превращается в инактивированную форму, при этом выделяются четыре атома водорода, которые используются в восстановительных реакциях, и два атома кислорода, поступающие в атмосферу.
Таким образом, первыми стабильными химическими продуктами световой реакции в растениях являются НАДФ — Н2 и АТФ.
В темновую фазу аминокислоты и белки образуются в цитоплазме.
Темновая фаза фотосинтеза служит продолжением световой фазы. В темновой фазе с участием АТФ и НАДФ — Н2 из углекислого газа строятся различные органические вещества.
При этом НАДФ — Н2 выполняет в темновой фазе роль восстановителя, а АТФ служит источником энергии. Восстановитель окисляется до НАДФ, а от АТФ отщепляется один остаток фосфорной кислоты (Н3РO4) и получается АДФ.
НАДФ и АДФ снова возвращаются из матрикса в граны, где в световой фазе снова преобразуются в НАДФ — Н2 и АТФ и все начинается сначала.
Последовательность реакций на пути превращения СO2 в сахар удалось выяснить благодаря применению радиоактивного углерода 14С. Было установлено, что в процессе фотосинтеза за несколько минут образуется большое число соединений.
Однако когда время, отведенное на фотосинтез, сократили до 0,5 с, удалось обнаружить лишь трехуглеродное фосфорилированное соединение — трифосфоглицериновую кислоту (ФГК). Следовательно, ФГК — это первый стабильный продукт, образующийся из СO2 в процессе фотосинтеза.
Оказалось, что первым веществом, которое соединяется с СO2 (акцептор СO2), является пятиуглеродное фосфорилированное соединение — рибулезодифосфат (РДФ), распадающееся после присоединения СO2 на две молекулы ФГК.
Фермент, катализирующий эту реакцию, — РДФ — карбоксилаза — занимает в количественном отношении первое место среди белков, содержащихся в белковой ткани.
Фосфоглицериновая кислота восстанавливается до уровня альдегида за счет восстановительного потенциала НАДФ — Н2 и энергии АТФ.
Фосфоглицериновый альдегид, представляющий собой фосфорилированное соединение сахара, содержит только три атома углерода, тогда как простейшие сахара содержат шесть атомов углерода. Для того чтобы образовалась гексоза (простейший сахар), две молекулы фосфоглицеринового альдегида должны соединиться и полученный продукт — гексозодифосфат — должен подвергнуться дефосфорилированию.
Получившаяся гексоза может направляться либо на синтез сахарозы и полисахаридов, либо на построение любых других органических соединений клетки. Таким образом, сахар, образующийся в процессе фотосинтеза из СO2, — это основное органическое вещество, которое в клетках высших растений служит источником как энергии, так и необходимых клетке строительных белков.
Углекислый газ. Интенсивность фотосинтеза зависит от количества углекислого газа в воздухе. Обычно в атмосферном воздухе содержится 0,03 % СO2. Увеличение его содержания способствует повышению урожайности, что используют при выращивании растений в парниках, оранжереях, теплицах. Установлено, что наилучшие условия для фотосинтеза создаются при содержании СO2 около 1,0%. Повышение содержания СO2 до 5,0% способствует повышению интенсивности фотосинтеза, но в этом случае необходимо повысить освещенность.
Количество СO2, усвоенное в единицу времени на единицу массы хлорофилла, называется ассимиляционным числом. Количество миллиграммов СO2, усвоенное за 1 ч на 1 дм2 листовой поверхности, называется интенсивностью фотосинтеза. Интенсивность фотосинтеза у различных видов растений неодинакова, изменяется она и с возрастом растений.
Свет. Растения поглощают 85 — 90 % попадающей на них световой энергии, но на фотосинтез идет только 1 — 5% от поглощенной световой энергии. Остальная энергия используется на нагрев растения и транспирацию. Все растения по их отношению к интенсивности освещения можно разделить на две группы — светолюбивые и тенелюбивые. Светолюбивые требуют большей освещенности, теневыносливые — меньшей. Вода. Обеспеченность растений водой имеет важное значение. Недостаточное насыщение клеток водой вызывает закрытие устьиц, а следовательно, снижает снабжение растений углекислым газом. Обезвоживание клеток нарушает деятельность ферментов. Температурный режим. Наилучший температурный режим для большинства растений, при котором фотосинтез идет наиболее интенсивно, 20 — 30 °С. При понижении или повышении температуры фотосинтез замедляется. Хлорофилл в клетках растений образуется при температуре от 2 до 40 °С. При благоприятном сочетании всех необходимых для фотосинтеза факторов растения наиболее активно накапливают органические вещества и выделяют кислород. Образующиеся в избытке продукты фотосинтеза — сахара — немедленно превращаются в высокополимерное запасное соединение — крахмал, откладывающийся в виде крахмальных зерен в хлоропластах и лейкопластах. Одновременно какая — то часть Сахаров выводится из пластид и перемещается в другие части растения. Крахмал может вновь расщепляться до Сахаров, которые, окисляясь в процессе дыхания, обеспечивают клетку энергией. Таким образом, искусственно регулируя газовый состав атмосферы, обеспечивая растения светом, водой, теплом, можно повышать интенсивность фотосинтеза и, следовательно, увеличивать продуктивность растений. Именно на это направлены агротехнические приемы при возделывании сельскохозяйственных культур: обогащение почвы органическими веществами, обработка почвы, орошение, мульчирование, регулирование густоты посевов и др.
читайте так-же
| Фотоплан | В началобуква «Ф»буквосочетание «ФО» | Фотосхема |
Статья «Фотосинтез» была прочитана 40654 раз
Источник: http://enc.sci-lib.com/article0001086.html
Механизм фотосинтеза. Световая и темновая стадии
В настоящее время установлено, что фотосинтез протекает в две стадии: световую и темновую. Световая стадия — это процесс использования света для расщепления воды; при этом выделяется кислород и образуются богатые энергией соединения.
Темновая стадия включает группу реакций, в которых используются высокоэнергетические продукты световой стадии для восстановления СО2 до простого сахара, т.е. для ассимиляции углерода. Поэтому темновую стадию называют также стадией синтеза.
Термин «темновая стадия» означает лишь то, что свет в ней непосредственно не участвует. Современные представления о механизме фотосинтеза сформировались на основе исследований, проведенных в 1930-1950-х годах.
До этого на протяжении многих лет ученых вводила в заблуждение на первый взгляд простая, однако неверная гипотеза, согласно которой О2 образуется из СО2, а освободившийся углерод реагирует с Н2О, в результате чего и образуются углеводы.
В 1930-х годах, когда выяснилось, что у некоторых серных бактерий кислород при фотосинтезе не выделяется, биохимик К. ван Ниль предположил, что кислород, выделяющийся в процессе фотосинтеза у зеленых растений, происходит из воды. У серных бактерий реакция протекает следующим образом:
Вместо О2 эти организмы образуют серу. Ван Ниль пришел к заключению, что все виды фотосинтеза можно описать уравнением
где Х — кислород в фотосинтезе, идущем с выделением О2, и сера в фотосинтезе серных бактерий. Ван Ниль также предположил, что этот процесс включает две стадии: световую и стадию синтеза. Эту гипотезу подкрепило открытие физиолога Р.Хилла.
Он обнаружил, что разрушенные или частично инактивированные клетки способны на свету осуществлять реакцию, в которой кислород выделяется, но СО2 не восстанавливается (ее назвали реакцией Хилла). Чтобы эта реакция могла идти, требовалось добавить какой-нибудь окислитель, способный присоединять электроны или водородные атомы, отдаваемые кислородом воды.
Один из реагентов Хилла — это хинон, который, присоединив два атома водорода, превращается в дигидрохинон. Другие реагенты Хилла содержали трехвалентное железо (ион Fe3+), которое, присоединив один электрон от кислорода воды, превращалось в двухвалентное (Fe2+).
Так было показано, что переход водородных атомов от кислорода воды на углерод может совершаться в форме независимого движения электронов и ионов водорода.
В настоящее время установлено, что для запасания энергии важен именно переход электронов от одного атома к другому, тогда как ионы водорода могут переходить в водный раствор, а при необходимости вновь из него извлекаться. Реакция Хилла, в которой световая энергия используется для того, чтобы вызвать перенос электронов от кислорода на окислитель (акцептор электронов), была первой демонстрацией перехода световой энергии в химическую и моделью световой стадии фотосинтеза.
Гипотеза, согласно которой кислород во время фотосинтеза непрерывно поступает от воды, нашла дальнейшее подтверждение в опытах с применением воды, меченной тяжелым изотопом кислорода (18О).
Поскольку изотопы кислорода (обычный 16О и тяжелый 18О) по своим химическим свойствам одинаковы, растения используют Н218О точно так же, как Н216О. Оказалось, что в выделенном кислороде присутствует 18О. В другом опыте растения вели фотосинтез с Н216О и С18О2.
При этом выделяемый в начале эксперимента кислород не содержал 18О.
В 1950-х годах физиолог растений Д.Арнон и другие исследователи доказали, что фотосинтез включает световую и темновую стадии. Из растительных клеток были получены препараты, способные осуществлять всю световую стадию.
Используя их, удалось установить, что на свету происходит перенос электронов от воды к фотосинтетическому окислителю, который в результате этого становится донором электронов для восстановления диоксида углерода на следующей стадии фотосинтеза. Переносчиком электронов служит никотинамидадениндинуклеотидфосфат.
Его окисленную форму обозначают НАДФ+, а восстановленную (образующуюся после присоединения двух электронов и иона водорода) — НАДФ?Н. В НАДФ+ атом азота пятивалентный (четыре связи и один положительный заряд), а в НАДФ?Н — трехвалентный (три связи). НАДФ+ принадлежит к т.н. коферментам.
Коферменты совместно с ферментами осуществляют многие химические реакции в живых системах, но в отличие от ферментов в ходе реакции изменяются. Бльшая часть преобразованной световой энергии, запасаемой в световой стадии фотосинтеза, запасается при переносе электронов от воды к НАДФ+.
Образовавшийся НАДФ?Н удерживает электроны не столь прочно, как кислород воды, и может отдавать их в процессах синтеза органических соединений, расходуя накопленную энергию на полезную химическую работу.
Значительное количество энергии запасается еще и другим способом, а именно в форме АТФ (аденозинтрифосфата).
Он образуется в результате отнятия воды от неорганического иона фосфата (HPO42-) и органического фосфата, аденозиндифосфата (АДФ), согласно следующему уравнению:
АТФ — богатое энергией соединение, и для его образования необходимо поступление энергии от какого-то источника. В обратной реакции, т.е. при расщеплении АТФ на АДФ и фосфат, энергия высвобождается.
Во многих случаях АТФ отдает свою энергию другим химическим соединениям в реакции, в которой водород замещается на фосфат.
В представленной ниже реакции сахар (ROH) фосфорилируется, превращаясь в сахарофосфат:
В сахарофосфате заключено больше энергии, чем в нефосфорилированном сахаре, поэтому его реакционная способность выше.
АТФ и НАДФ?Н, образующиеся (наряду с О2) в световой стадии фотосинтеза, используются затем на стадии синтеза углеводов и других органических соединений из диоксида углерода.
Источник: https://vuzlit.ru/2267407/mehanizm_fotosinteza_svetovaya_temnovaya_stadii
Фотосинтез и его фазы (световая и темновая)
Фотосинтез — уникальная система процессов создания с помощью хлорофилла и энергии света органических веществ из неорганических и выделения кислорода в атмосферу, реализуемая в огромных масштабах на суше и в воде.
Фотосинтез происходит в клетках зелёных растений с помощью пигментов, главным образом хлорофилла, находящегося в хлоропластах клетки. Его продуктами являются мономеры углеводов (моносахариды: глюкоза, фруктоза и др.).
В основе фотосинтеза лежит окислительно-восстановительный процесс, в котором электроны переносятся от донора-восстановителя (вода, водород и др.) к акцептору (CO2 , ацетат и др.) с образованием восстановленных соединений (углевода) и выделением кислорода, если окисляется вода. Фотосинтезирующие бактерии часто используют другие доноры, а не воду, кислород при этом они не выделяют.
В системе процессов фотосинтеза различают два цикла реакций, как две фазы, последовательно и непрерывно идущие друг за другом — световую и темновую (рис. 62).
Световая фаза фотосинтеза характеризуется тем, что здесь все процессы происходят только при участии энергии света, поэтому её и называют световой. Связывание солнечной (электромагнитной) энергии происходит преимущественно на мембранах тилакоидов хлоропласта. Размещающийся здесь хлорофилл и другие пигменты собраны в функциональные единицы-комплексы — пигментные системы, получившие название фотосистемы.
| Рис. 62. Схема фотосинтеза |
Таким образом, светособирающие и пигментно-белковые комплексы фотосистемы I и фотосистемы II обеспечивают процесс фотосинтеза необходимой энергией в виде макроэнергетических соединений НАДФ•Н и АТФ. В этом заключается основная функция световой фазы фотосинтеза. Она реализуется только при участии света и с помощью пигментов, размещённых в тилакоидной мембране хлоропластов.
Темновая фаза фотосинтеза проходит в строме хлоропласта без непосредственного поглощения света, в любое время суток. В процессе световой фазы фотосинтеза накапливается достаточно высокий уровень АТФ и НАДФ•Н. Однако сами по себе эти высокоэнергетические соединения не способны синтезировать углеводы из CO2. Поэтому становится очевидным, что и темновая фаза фотосинтеза — сложный процесс, включающий большое количество реакций. Материал с сайта http://doklad-referat.ru
Все процессы темновой фазы фотосинтеза идут без непосредственного потребления света, но в них большую роль играют высокоэнергетические вещества (АТФ и НАДФ•Н), образующиеся с участием энергии света, во время световой фазы фотосинтеза. В процессе темновой фазы энергия макроэнергетических связей АТФ преобразуется в химическую энергию органических соединений молекул углеводов. Это значит, что энергия солнечного света как бы консервируется в химических связях между атомами органических веществ, что имеет огромное значение в энергетике биосферы и конкретно для жизнедеятельности всего живого населения нашей планеты.
Фотосинтез происходит в хлоропластах клетки и представляет собой синтез углеводов в хлорофиллоносных клетках, идущий с потреблением энергии солнечного света. Различают световую и темповую фазы фотосинтеза. Световая фаза при непосредственном потреблении квантов света обеспечивает процесс синтеза необходимой энергией в виде НАДН и АТФ. Темновая фаза — без участия света, но путем многочисленного ряда химических реакций (цикл Кальвина) обеспечивает образование углеводов, главным образом глюкозы. Значение фотосинтеза в биосфере огромно.
На этой странице материал по темам:
Реферат темновая фаза фотосинтеза
Фотосинтез световая и темновая фазы реферат
План конспект скачать бесплатно процесс фотосинтеза и его фазы
Световая стадии фотосинтеза лекции
Фотосинтез темновая и световая фаза кратко
Вопросы по этому материалу:
Где происходит фотосинтез в растительной клетке?
Какие превращения происходят во время световой фазы фотосинтеза?
Что является результатом световой фазы фотосинтеза?
Где протекают темновые реакции фотосинтеза?
Природные солнечные батареи – Наука – Коммерсантъ
На Земле работают природные химические фабрики, в колоссальных количествах производящие химические вещества за счет энергии света — фотосинтезирующие организмы. Растения и фитопланктон поглощают 300 млрд тонн углекислого газа ежегодно, выделяя при этом 200 млрд тонн кислорода.
И это лишь оксигенирующие организмы — те, что в ходе эволюции научились окислять воду, выделяя из нее кислород. Более эволюционно ранние организмы такие превращения осуществлять не могут и живут за счет окисления других веществ, например сероводорода или соединений железа. Строение фотосинтезирующих организмов и условия их обитания очень разнообразны, но принципы, на основе которых они используют свет для синтеза химических веществ, во многом похожи. Изучение этих принципов позволяет лучше понять то, как устроена живая природа, а их воплощение на практике — создать искусственные преобразователи световой энергии.
Фотосинтез — сложный многостадийный процесс, который можно разделить на две фазы — световую и темновую. Световая фаза состоит в использовании энергии света для синтеза универсальной энергетической «валюты» живых клеток — аденозинтрифосфата (АТФ) и никотинамидадениндинуклеотидфосфата (НАДФН). Во время темновой фазы запасенная в этих веществах энергия расходуется на синтез углеводов и других органических веществ. Несмотря на название, темновая фаза может протекать и днем и ночью. Световая же фаза возможна лишь при освещении и протекает в специализированных комплексах органических пигментов с белком, которые называются фотосистемами.
Фотосистемы — сложноорганизованные структуры, находящиеся в клеточных (у некоторых бактерий) или внутриклеточных (у растений) мембранах. Они очень малы — всего несколько нанометров? но по химическим меркам они огромны и состоят из сотен тысяч атомов. Функционально фотосистемы состоят из двух типов частей — фотоантенн и реакционного центра. Задача фотоантенн — уловить квант света и передать его энергию в виде порции электронного возбуждения (так называемого экситона) реакционному центру. Попавший в реакционный центр экситон вызывает перенос электрона из одной его части в другую, что приводит к возникновению электрического потенциала по разные стороны мембраны, в котором находится фотосистема. Этот потенциал и является той движущей силой, которая вызывает химические превращения.
Поскольку даже в яркий солнечный день фотоны попадают на фотосистему не так уж часто, фотоантенны составляют ее большую часть. Каждая из них содержит десятки, а иногда и сотни молекул пигментов — зеленых хлорофиллов и желто-оранжево-красных каротиноидов, удерживаемых друг относительно друга в определенном положении белком. На один реакционный центр приходится несколько фотоантенн, и их количество может меняться — так фотосинтезирующие организмы приспосабливаются к разным условиям освещенности. Несмотря на внушительные размеры фотосистем, экситон, возникший после поглощения света в любой из фотоантенн, с вероятностью более 90% дойдет до реакционного центра, и притом за очень короткое время — за 10–12–10–10 секунд. Как ему это удается, учитывая, что состояние электронного возбуждения очень нестабильно и склонно к быстрой релаксации, то есть полному рассеиванию своей энергии?
Оказывается, белок в фотосистемах выполняет не только структурную роль. За счет специфических взаимодействий белок меняет энергию возбуждения хлорофиллов таким образом, что она немного понижается по мере приближения к реакционному центру. Благодаря этому движение экситона в фотосистеме больше похоже на течение реки, чем на случайные блуждания: он все время перемещается в сторону молекул с меньшей энергией возбуждения и рано или поздно приходит к той ее части, где эта энергия наименьшая,— реакционному центру. При этом на каждом шаге часть энергии экситона рассеивается в белке. Такой механизм обеспечивает направленность переноса и его высокую скорость. Однако у этого механизма есть цена: энергия, дошедшая до реакционного центра меньше, чем та, которой изначально обладал поглощенный фотон. Белок также выполняет и защитную функцию, служа своего рода термостатом: в том случае, если экситон все-таки не дошел реакционного центра, его энергия рассеивается по белковой молекуле, что предотвращает повреждение пигментов фотосистемы.
Из-за больших размеров фотосистемы непросто выделять и изучать на практике. Впервые трехмерная структура компонентов фотосистем с атомным разрешением была установлена лишь в 1990-х годах, и примерно в это же время появилась техническая возможность изучения динамики экситона в них спектроскопическими методами. В этой ситуации полезную информацию дают теоретические методы исследования.
На химическом факультете МГУ ведутся работы по моделированию работы природных фотосистем и разработке их искусственных аналогов. Для описания работы таких объектов и предсказания их эффективности необходим учет множества факторов, включая взаимное расположение пигментов и энергию их электростатического взаимодействия, свойства среды, в которой они находятся, температуру и многих других. Также разными могут быть и желаемые параметры функционирования: для аккумулирования энергии необходимы ее малые потери в среде при переносе экситона, а если последний служит переносчиком сигнала, то важнее становятся скорость переноса и малая вероятность релаксации.
Распределение заряда в молекуле хлорофилла b
Описание фотосистем и процессов, составляющих световую фазу фотосинтеза, возможно только на основе квантовой механики. Одно из ключевых отличий квантовой механики от классической, с проявлениями которой мы сталкиваемся в быту, является то, что у квантовой системы возможны не любые энергетические состояния, а только определенные, или, как говорят, дискретные. Соответственно, и переходы между этими состояниями также сопровождаются испусканием или поглощением строго определенного количества энергии. Это справедливо и для пигментов (благодаря чему каждый из них имеет определенный цвет), и для белка, колебательные состояния которого также квантуются. Квантовомеханический анализ работы фотосистем показывает, что структура белка в них тонко подстроена под остальные компоненты. Каждый переход экситона от пигмента к пигменту сопровождается рассеянием небольшого количества энергии, которая в точности достаточна для перевода близлежащих атомов белка на один колебательный уровень выше. За счет этого достигается высокая скорость переноса экситона. Этот механизм был выработан фотосинтезирующими организмами в ходе эволюции. Подобная структурная сложность среды, в которую погружены пигменты, пока недостижима для искусственных фотосистем, поэтому они уступают по характеристикам своим природным прототипам.
Александр Белов, химический факультет МГУ им. М. В. Ломоносова
Фотосинтез
Задачи: Сформировать знания о реакциях пластического и энергетического обменов и их взаимосвязи; вспомнить особенности строения хлоропластов. Дать характеристику световой и темновой фазы фотосинтеза. Показать значение фотосинтеза как процесса, обеспечивающего синтез органических веществ, поглощение углекислого газа и выделение кислорода в атмосферу.
Тип урока: лекция.
Оборудование:
- Средства наглядности: таблицы по общей биологии;
- ТСО: компьютер; мультимедиапроектор.
План лекции:
- История изучения процесса.
- Эксперименты по фотосинтезу.
- Фотосинтез, как анаболический процесс.
- Хлорофилл и его свойства.
- Фотосистемы.
- Световая фаза фотосинтеза.
- Темновая фаза фотосинтеза.
- Лимитирующие факторы фотосинтеза.
Ход лекции
История изучения фотосинтеза
1630 год начало изучения фотосинтеза. Ван Гельмонт доказал, что растения образуют органические вещества, а не получают их из почвы. Взвешивая горшок с землей и ивой, и отдельно само дерево, он показал, что через 5 лет масса дерева увеличилась на 74 кг, тогда как почва потеряла только 57 г. Он решил, что пищу дерево получает из воды. В настоящее время мы знаем, что используется углекислый газ.
В 1804 году Соссюр установил, что в процессе фотосинтеза велико значение воды.
В 1887 году открыты хемосинтезирующие бактерии.
В 1905 году Блэкман установил, что фотосинтез состоит из двух фаз: быстрой – световой и ряда последовательных медленных реакций темновой фазы.
Эксперименты по фотосинтезу
| 1 опыт доказывает значение солнечного света (рис. 1.) | 2 опыт доказывает значение углекислого газа для фотосинтеза (рис. 2.) |
3 опыт доказывает значение фотосинтеза (рис.3.) | |
| |
Фотосинтез, как анаболический процесс
- Ежегодно в результате фотосинтеза образуется 150 млрд. тонн органического вещества и 200 млрд. тонн свободного кислорода.
- Круговорот кислорода, углерода и других элементов, вовлекаемых в фотосинтез. Поддерживает современный состав атмосферы, необходимый для существования современных форм жизни.
- Фотосинтез препятствует увеличению концентрации углекислого газа, предотвращая перегрев Земли вследствие парникового эффекта.
- Фотосинтез – основа всех цепей питания на Земле.
- Запасенная в продуктах энергия – основной источник энергии для человечества.
Сущность фотосинтеза заключается в превращении световой энергии солнечного луча в химическую энергию в виде АТФ и НАДФ·Н2.
Суммарное уравнение фотосинтеза:
6СО2 + 6Н2О → С6Н12О6 + 6О2↑
Существует два главных типа фотосинтеза:
| анаэробный | аэробный |
| Характерен для фотосинтезирующих бактерий (подцарство Настоящие бактерии). Фотосинтезирующим пигментом у них является бактериохлорофилл. Кислород не выделяется. | Характерен для всех оксифотобактерий и зеленых растений. Фотосинтез в растениях осуществляется в хлоропластах содержащих хлорофилл. Кислород выделяется. |
Хлорофилл и его свойства
|
Рис.4. Структурная формула хлорофилла а | Молекула хлорофилла имеет эмпирическую формулу: С55Н72О5N4Мg. Атомы С, Н, О, N соединены в сложное порфириновое кольцо. Хлорофилл близок по строению к гемоглобину крови, только в гемме в центре молекулы атом Fe, а в хлорофилле атом Мg, связанный с одним или четырьмя атомами азота. Молекула хлорофилла имеет длинный «хвост» — остаток спирта фитола, который содержит цепь из 20 углеродных атомов. |
Виды хлорофилла
Хлорофилл имеет модификации а, в, с, d. Отличаются они структурным строением и спектром поглощения света. Например: хлорофилл в содержит на один атом кислорода больше и на два атома водорода меньше, чем хлорофилл а.
Все растения и оксифотобактерии имеют как основной пигмент желто-зеленый хлорофилл а, а как дополнительный хлорофилл в.
Другие пигменты растений
Некоторые другие пигменты способны поглощать солнечную энергию и передавать ее в хлорофилл, вовлекая ее тем самым в фотосинтез.
У большинства растений есть темно оранжевый пигмент – каротин, который в животном организме превращается в витамин А и желтый пигмент – ксантофилл.
Фикоцианин и фикоэритрин – содержат красные и сине-зеленые водоросли. У красных водорослей эти пигменты принимают более активное участие в процессе фотосинтеза, чем хлорофилл.
Хлорофилл минимально поглощает свет в сине-зеленой части спектра. Хлорофилл а, в- в фиолетовой области спектра, где длина волны 440 нм. Уникальная функция хлорофилла состоит в том, что он интенсивно поглощает солнечную энергию и передает ее другим молекулам.
Пигменты поглощают определенную длину волны, не поглощенные участки солнечного спектра отражаются, что обеспечивает окраску пигмента. Зеленый свет не поглощается, поэтому хлорофилл зеленый.
Пигменты – это химические соединения, которые поглощают видимый свет, что приводит электроны в возбужденное состояние. Чем меньше длина волны, тем больше энергия света и больше его способность переводить электроны в возбужденное состояние. Это состояние неустойчиво и вскоре вся молекула возвращается в свое обычное низкоэнергетическое состояние теряя при этом энергию возбуждения. Эта энергия может быть использована на флуоресценцию.
Фотосистемы
Пигменты растений участвующие в фотосинтезе «упакованы» в тилакоиды хлоропластов в виде функциональных фотосинтетических единиц – фотосинтетических систем: фотосистемы I и фотосистемы II.
Каждая система состоит из набора вспомогательных пигментов (от 250 до 400 молекул), передающих энергию на одну молекулу главного пигмента и она называется реакционным центром. В нем энергия Солнца используется для фотохимических реакций.
|
Рис.5. Фотосистемы | Фотосистема I имеет более мелкие частицы, чем фотосистема II. Частицы фотосистемы II связаны с гранами. Энергия захватывается как бы в ловушку со вспомогательных (антенных) пигментов на главный. Это может быть хлорофилл а – Р690 или Р700 (Р – пигмент, а 690-700 – максимально поглощенная длина волны в нм). Р690 и Р700 – энергетические ловушки |
| Фотосинтез | ||
| Световая фаза Фотофизический этап | Световая фаза Фотохимический этап | Темновая фаза или цикл Кальвина |
| Поглощение квантов света пигментами, идет возбуждение электронов в этих молекулах и передача возбуждения от одной молекулы к другой. | Преобразование энергии света в энергию химических связей АТФ и НАДФ.Н2. Идет в фотосинтетических мембранах. | Идет за счет энергии, которая образовалась в световой фазе. Суть процесса: включение углекислого газа в образование органических веществ. |
Световая фаза идет обязательно с участием света, темновая фаза и на свету и в темноте. Световой процесс происходит в тилакоидах хлоропластов, темновой – в строме, т.е. эти процессы пространственно разобщены.
Световая фаза фотосинтеза
В 1958 году Арнон и его сотрудники изучили световую фазу фотосинтеза. Они установили, что источником энергии при фотосинтезе является свет, а так как на свету в хлорофилле происходит синтез из АДФ+Ф.к. → АТФ, то этот процесс называется фосфорилированием. Оно сопряжено с переносом электронов в мембранах.
Роль световых реакций: 1. Синтез АТФ – фосфорилирование. 2. Синтез НАДФ.Н2.
Путь переноса электронов называется Z-схемой.
Z-схема. Нециклическое и циклическое фотофосфорилирование (рис. 6.)
-
Начало процесса. Поглощение квантов света. Квант света попадает на ФС II, находящуюся в мембранах тилакоидов гран и приводит к возбуждению пигментов – это возбуждение передается от одной молекулы антенного пигмента к другой вплоть до реакционного центра. Все электроны собираются вокруг ловушки и отдают энергию виде электронов в электроннотранспортную цепь. Электрон, поглотив фотон, отрывается от молекулы хлорофилла и переходит на более высокий энергетический уровень присоединяясь к молекулам-переносчикам. Затем он двигается по электроннотранспортной цепи переходя от одного переносчика к другому (от пластохинона к пластоцианину) постепенно растрачивая энергию. Часть этой энергии растрачивается на синтез АТФ.
-
Нециклическое фотофосфорилирование. Растратив энергию электрон достигает ФСI, где он опять поглощает фотон и снова поднимается еще на более высокий энергетический уровень, и пройдя через несколько переносчиков (ферредоксин) передается конечному акцептору цепи НАДФ+, который расположен на внешней стороне мембраны тилакоида.
-
Фоторазложение или фотолиз воды. Поглотив фотон электроны отрываются от молекул хлорофилла реакционного центра ФС II и через ФС I переходят к НАДФ+. Пока на место ушедшего электрона в ФС II не встанет другой, она не сможет функционировать. Место ушедших электронов занимают электроны воды, которая находится во внутреннем пространстве тилакоида. При этом происходит светозависимое разложение воды или фотолиз («фото» — свет): Н2О →2 Н+ 2е + ½ О2. При фотолизе вода распадается на протоны, электроны и кислород. Процесс происходит с участием ферментов локализованных на внутренней мембране тилакоидов. Образовавшийся кислород выделяется в окружающую среду. Протоны накапливаются во внутреннем пространстве тилакоидов, образуя резервуар протонов. Таки образом при нециклическом потоке электронов от ФС II к НАДФ+ в конечном счете транспортируются электроны воды.
-
Процесс химио-осмоса.
Рис.7. Процесс фотосинтеза
- Свет, попадая на молекулы хлорофилла, которые находятся в мембранах тилакоидов гран, приводит их в возбужденное состояние. В результате этого электроны сходят со своих орбит и переносятся с помощью переносчиков за пределы мембраны тилакоида, где накапливаются, создавая отрицательно заряженное электрическое поле.
- Протоны, образовавшиеся при фотолизе, не проникают через мембрану тилакоида и накапливаются внутри, образуя положительно заряженное электрическое поле, что приводит к увеличению разности потенциалов по обе стороны мембраны.
- При достижении критической разности потенциалов, протоны могут выходить в строму по протонному каналу. С каналами связаны ферменты АТФ-синтетазы, которые используют энергию протонов на синтез АТФ. На каждые три протона, которые проходят через канал, синтезируется одна молекула АТФ. Большая часть АТФ при фотосинтезе образуется этим путем.
- Протоны, вышедшие на поверхность мембраны тилакоида, соединяются с электронами, образуя атомарный водород, который идет на восстановление переносчика НАДФ+.
-
Циклический поток электронов. ФС I может работать независимо от ФСII. Под действием света, электрон выбивается из молекулы хлорофилла реакционного центра ФС I, передается к тому же акцептору, что и при нециклическом потоке, но далее идет не к НАДФ+, а по обходному пути возвращается на тоже место в ФС I. Поглощенная электроном энергия используется на синтез АТФ. Таким образом электрон двигается по кольцу. Это циклический поток.
В ходе циклического транспорта электронов не происходит образования НАДФ.Н2 и фоторазложения Н2О, следовательно и выделение О2. Этот путь используется тогда, когда в клетке избыток НАДФ.Н2, но требуется дополнительная АТФ.
Все эти процессы относятся к световой фазе фотосинтеза. В дальнейшем энергия АТФ и НАДФ.Н2 используется для синтеза глюкозы. Для этого процесса свет не нужен. Это реакции темновой фазы фотосинтеза.
Темновая фаза фотосинтеза или цикл Кальвина
Синтез глюкозы происходит в ходе циклического процесса, который получил название по имени ученого Мельвина Кальвина, открывшего его, и награжденного Нобелевской премией.
Рис. 8. Цикл Кальвина
Каждая реакция цикла Кальвина осуществляется своим ферментом. Для образования глюкозы используются: СО2, протоны и электроны от НАДФ.Н2, энергия АТФ и НАДФ.Н2. Происходит процесс в строме хлоропласта. Исходным и конечным соединением цикла Кальвина, к которому с помощью фермента рибулозодифосфаткарбоксилазы присоединяется СО2, является пятиуглеродный сахар – рибулозобифосфат, содержащий две фосфатные группы. В результате образуется шестиуглеродное соединение, сразу же распадающееся на две трехуглеродные молекулы фосфоглицериновой кислоты, которые затем восстанавливаются до фосфоглицеринового альдегида. При этом, часть образовавшегося фосфоглицеринового альдегида используется для регенерации рибулозобифосфата, и, таким образом, цикл возобновляется снова (5С3 → 3С5), а часть используется для синтеза глюкозы и других органических соединений (2С3 → С6 → С6Н12О6).
Для образования одной молекулы глюкозы необходимо 6 оборотов цикла и требуется 12НАДФ.Н2 и 18 АТФ. Из суммарного уравнения реакции получается:
6СО2 + 6Н2О → С6Н12О6 + 6О2
Из приведенного уравнения видно, что атомы С и О вошли в глюкозу из СО2, а атомы водорода из Н2О. Глюкоза в дальнейшем может быть использована как на синтез сложных углеводов (целлюлозы, крахмала), так и на образование белков и липидов.
(С4 – фотосинтез. В 1965 году было доказано, что у сахарного тростника – первыми продуктами фотосинтеза, являются кислоты, содержащие четыре атома углерода (яблочная, щавелевоуксусная, аспарагиновая). К С4 растениям принадлежат кукуруза, сорго, просо).
Лимитирующие факторы фотосинтеза
Скорость фотосинтеза – наиболее важный фактор влияющий на урожайность с/х культур. Так, для темновых фаз фотосинтеза нужны НАДФ.Н2 и АТФ, и поэтому скорость темновых реакций зависит от световых реакций. При слабой освещенности скорость образования органических веществ будет мала. Поэтому свет – лимитирующий фактор.
Из всех факторов одновременно влияющих на процесс фотосинтеза лимитирующим будет тот, который ближе к минимальному уровню. Это установил Блэкман в 1905 году. Разные факторы могут быть лимитными, но один из них главный.
- При низкой освещенности скорость фотосинтеза прямопропорциональна интенсивности света. Свет – лимитирующий фактор при низкой освещенности. При большой интенсивности света происходит обесцвечивание хлорофилла и фотосинтез замедляется. В таких условиях в природе растения обычно защищены (толстая кутикула, опушенные листья, чешуйки).
- Для темновых реакций фотосинтеза необходим углекислый газ, который включается в органические вещества, в полевых условиях является лимитирующим фактором. Концентрация СО2 варьирует в атмосфере в пределах от 0,03–0,04%, но если повысить ее, то можно увеличить скорость фотосинтеза. Некоторые тепличные культуры сейчас выращиваются при повышенном содержании СО2.
- Температурный фактор. Темновые и некоторые световые реакции фотосинтеза контролируются ферментами, а их действие зависит от температуры. Оптимальная температура для растений умеренного пояса составляет 25 °С. При каждом повышении температуры на 10 °С (вплоть до 35 °С) скорость реакций удваивается, но из-за влияния ряда иных факторов растения лучше растут при 25 °С.
- Вода – исходное вещество для фотосинтеза. Недостаток воды влияет на многие процессы в клетках. Но даже временное увядание приводит к серьезным потерям урожая. Причины: при увядании устьица растений закрываются, а это мешает свободному доступу СО2 для фотосинтеза; при нехватке воды в листьях некоторых растений накапливается абсцизовая кислота. Это гормон растений – ингибитор роста. В лабораторных условиях ее используют для изучения торможения ростового процесса.
- Концентрация хлорофилла. Количество хлорофилла может уменьшаться при заболеваниях мучнистой росой, ржавчиной, вирусными болезнями, недостатком минеральных веществ и возрастом (при нормальном старении). При пожелтении листьев наблюдаются хлоротичные явления или хлороз. Причиной может быть недостаток минеральных веществ. Для синтеза хлорофилла нужны Fe, Mg, N и К.
- Кислород. Высокая концентрация кислорода в атмосфере (21%) ингибирует фотосинтез. Кислород конкурирует с углекислым газом за активный центр фермента, участвующего в фиксации СО2, что снижает скорость фотосинтеза.
- Специфические ингибиторы. Лучший способ погубить растение – это подавить фотосинтез. Для этого ученые разработали ингибиторы – гербициды – диоксины. Например: ДХММ – дихлорфенилдиметилмочевина – подавляет световые реакции фотосинтеза. Успешно используют для изучения световых реакций фотосинтеза.
- Загрязнение окружающей среды. Газы промышленного происхождения, озон и сернистый газ, даже в малых концентрациях сильно повреждают листья у ряда растений. К сернистому газу очень чувствительны лишайники. Поэтому существует метод лихеноиндикации – определение загрязнения окружающей среды по лишайникам. Сажа забивает устьица и уменьшает прозрачность листовой эпидермы, что снижает скорость фотосинтеза.
Космическая роль растений (описана К. А. Тимирязевым) заключается в том, что растения – единственные организмы, усваивающие солнечную энергию и аккумулирующие ее в виде потенциальной химической энергии органических соединений. Выделяющийся О2 поддерживает жизнедеятельность всех аэробных организмов. Из кислорода образуется озон, который защищает все живое от ультрафиолетовых лучей. Растения использовали из атмосферы громадное количество СО2, избыток которого создавал «парниковый эффект», и температура планеты понизилась до нынешних значений.
Две стадии фотосинтеза | Sciencing
Фотосинтез представляет собой биологический процесс, с помощью которого растения преобразуют световую энергию в сахар для топливных клеток растений. Состоит из двух стадий, одна стадия превращает световую энергию в сахар, а затем клеточное дыхание превращает сахар в аденозинтрифосфат, известный как АТФ, топливо для всей клеточной жизни. Преобразование неиспользуемого солнечного света делает растения зелеными.
Хотя механизмы фотосинтеза сложны, общая реакция протекает следующим образом: углекислый газ + солнечный свет + вода —> глюкоза (сахар) + молекулярный кислород.Фотосинтез проходит в несколько этапов, которые происходят на двух этапах: светлая фаза и темная фаза.
Этап первый: световые реакции
В светозависимом процессе, который происходит в гране, многослойной мембранной структуре внутри хлоропластов, прямая энергия света помогает растению производить молекулы, которые несут энергию для использования в темной фазе. фотосинтеза. Растение использует световую энергию для выработки кофермента никотинамидадениндинуклеотидфосфата, или НАДФН и АТФ, молекул, переносящих энергию.Химические связи в этих соединениях хранят энергию и используются во время темной фазы.
Вторая стадия: темные реакции
Темная фаза, которая имеет место в строме и в темноте, когда присутствуют молекулы, несущие энергию, также известна как цикл Кальвина или цикл C 3 . Темная фаза использует АТФ и НАДФН, образующиеся в светлой фазе, для образования ковалентных связей C-C углеводов из диоксида углерода и воды с химическим бифосфатом рибулозы или RuBP, химическим веществом 5-C, улавливающим диоксид углерода.В цикл вступают шесть молекул углекислого газа, который, в свою очередь, производит одну молекулу глюкозы или сахара.
Как работает фотосинтез
Ключевым компонентом, который управляет фотосинтезом, является молекула хлорофилла. Хлорофилл — это большая молекула со специальной структурой, которая позволяет ему захватывать световую энергию и преобразовывать ее в электроны с высокой энергией, которые используются во время реакций двух фаз для получения сахара или глюкозы.
У фотосинтезирующих бактерий реакция происходит в клеточной мембране и внутри клетки, но вне ядра.У растений и фотосинтезирующих простейших — простейшие — это одноклеточные организмы, принадлежащие к области эукариот, той же области жизни, которая включает растения, животных и грибы — фотосинтез происходит в хлоропластах. Хлоропласты представляют собой тип органелл или мембраносвязанных компартментов, адаптированных для определенных функций, таких как создание энергии для растений.
Хлоропласты — история эволюции
Хотя сегодня хлоропласты существуют в других клетках, таких как клетки растений, у них есть собственная ДНК и гены.Анализ последовательности этих генов показал, что хлоропласты произошли от независимо живущих фотосинтезирующих организмов, связанных с группой бактерий, называемых цианобактериями.
Подобный процесс произошел, когда предки митохондрий, органелл внутри клеток, где происходит окислительное дыхание, химическая противоположность фотосинтезу. Согласно теории эндосимбиоза, теории, получившей развитие недавно благодаря новому исследованию, опубликованному в журнале Nature, и хлоропласты, и митохондрии когда-то жили как независимые бактерии, но были поглощены предками эукариот, что в конечном итоге привело к появление растений и животных.
Фотосинтез | Биология для майоров I
Определите основные компоненты и этапы фотосинтеза
Процессы во всех организмах — от бактерий до людей — требуют энергии. Чтобы получить эту энергию, многие организмы получают доступ к накопленной энергии, поедая, то есть глотая другие организмы. Но откуда берется энергия, накопленная в пище? Вся эта энергия восходит к фотосинтезу.
Фотосинтез необходим для всей жизни на Земле; от него зависят и растения, и животные.Это единственный биологический процесс, который может улавливать энергию, исходящую из космоса (солнечный свет), и преобразовывать ее в химические соединения (углеводы), которые каждый организм использует для обеспечения своего метаболизма. Короче говоря, энергия солнечного света улавливается и используется для возбуждения электронов, которые затем сохраняются в ковалентных связях молекул сахара. Насколько долговечны и стабильны эти ковалентные связи? Энергия, извлекаемая сегодня при сжигании угля и нефтепродуктов, представляет собой энергию солнечного света, захваченную и сохраненную в процессе фотосинтеза около 300 миллионов лет назад.
Рис. 1. Фотоавтотрофы, включая (а) растения, (б) водоросли и (в) цианобактерии, синтезируют свои органические соединения посредством фотосинтеза, используя солнечный свет в качестве источника энергии. Цианобактерии и планктонные водоросли могут расти на огромных участках воды, иногда полностью покрывая поверхность. В (d) глубоководном источнике хемоавтотрофы, такие как эти (e) термофильные бактерии, улавливают энергию неорганических соединений для производства органических соединений. В экосистеме, окружающей вентиляционные отверстия, обитает множество разнообразных животных, таких как трубчатые черви, ракообразные и осьминоги, которые получают энергию от бактерий.(кредит a: модификация работы Стива Хиллебранда, Служба охраны рыбных ресурсов и дикой природы США; кредит b: модификация работы с помощью «эвтрофикации и гипоксии» / Flickr; кредит c: модификация работы НАСА; кредит d: Вашингтонский университет, NOAA; кредит e : модификация работы Марка Аменда, Центр подводных исследований западного побережья и полярных регионов, UAF, NOAA)
Рис. 2. Энергия, запасенная в молекулах углеводов в результате фотосинтеза, проходит через пищевую цепь. Хищник, поедающий этих оленей, получает часть энергии, происходящей от фотосинтезирующей растительности, которую олени потребляли.(кредит: модификация работы Стива Ван Рипера, Служба охраны рыболовства и дикой природы США)
Растения, водоросли и группа бактерий, называемых цианобактериями, — единственные организмы, способные к фотосинтезу (рис. 1). Поскольку они используют свет для производства собственной пищи, их называют фотоавтотрофами (буквально «самокормящие с помощью света»). Другие организмы, такие как животные, грибы и большинство других бактерий, называются гетеротрофами («другие кормушки»), потому что они должны полагаться на сахара, производимые фотосинтезирующими организмами, для удовлетворения своих энергетических потребностей.Третья очень интересная группа бактерий синтезирует сахар, не используя энергию солнечного света, а извлекая энергию из неорганических химических соединений; следовательно, их называют хемоавтотрофами .
Важность фотосинтеза заключается не только в том, что он может улавливать энергию солнечного света. Ящерица, загорающая в холодный день, может использовать солнечную энергию, чтобы согреться. Фотосинтез жизненно важен, потому что он развился как способ хранения энергии солнечного излучения (часть «фото») в виде высокоэнергетических электронов в углерод-углеродных связях молекул углеводов (часть «синтез»).Эти углеводы являются источником энергии, который гетеротрофы используют для синтеза АТФ посредством дыхания. Таким образом, фотосинтез обеспечивает работу 99 процентов экосистем Земли. Когда высший хищник, такой как волк, охотится на оленя (рис. 2), волк находится в конце энергетического пути, который перешел от ядерных реакций на поверхности солнца к свету, фотосинтезу и растительности, оленям и, наконец, волкам.
Цели обучения
- Определить реагенты и продукты фотосинтеза
- Опишите видимый и электромагнитный спектры света применительно к фотосинтезу.
- Опишите светозависимые реакции, происходящие во время фотосинтеза.
- Определите светонезависимые реакции фотосинтеза
Фотосинтез — это многоступенчатый процесс, требующий солнечного света, двуокиси углерода (с низким содержанием энергии) и воды в качестве субстратов (рис. 3).После завершения процесса он высвобождает кислород и производит глицеральдегид-3-фосфат (GA3P), простые молекулы углеводов (с высоким содержанием энергии), которые впоследствии могут быть преобразованы в глюкозу, сахарозу или любую из десятков других молекул сахара. Эти молекулы сахара содержат энергию и активированный углерод, необходимый всем живым существам для выживания.
Рис. 3. Фотосинтез использует солнечную энергию, углекислый газ и воду для производства энергоемких углеводов. Кислород образуется как побочный продукт фотосинтеза.
Ниже приводится химическое уравнение фотосинтеза (рис. 4):
Рис. 4. Основное уравнение фотосинтеза обманчиво просто. В действительности процесс проходит в несколько этапов с участием промежуточных реагентов и продуктов. Глюкоза, основной источник энергии в клетках, состоит из двух трехуглеродных GA3P.
Хотя уравнение выглядит простым, многие этапы фотосинтеза на самом деле довольно сложны. Прежде чем изучать детали того, как фотоавтотрофы превращают солнечный свет в пищу, важно ознакомиться с соответствующими структурами.
У растений фотосинтез обычно происходит в листьях, которые состоят из нескольких слоев клеток. Процесс фотосинтеза происходит в среднем слое, называемом мезофиллом . Газообмен углекислого газа и кислорода происходит через небольшие регулируемые отверстия, называемые устьицами (единственное число: стома), которые также играют роль в регулировании газообмена и водного баланса. Устьица обычно расположены на нижней стороне листа, что помогает свести к минимуму потерю воды.Каждая стома окружена замыкающими клетками, которые регулируют открытие и закрытие устьиц, набухая или сжимаясь в ответ на осмотические изменения.
У всех автотрофных эукариот фотосинтез происходит внутри органеллы, называемой хлоропластом . У растений хлоропластсодержащие клетки существуют в мезофилле. Хлоропласты имеют двойную мембранную оболочку (состоящую из внешней и внутренней мембран). Внутри хлоропласта расположены многослойные дискообразные структуры, называемые тилакоидами .В тилакоидную мембрану встроен хлорофилл, пигмент , (молекула, поглощающая свет), ответственный за начальное взаимодействие между светом и растительным материалом, и многочисленные белки, составляющие цепь переноса электронов. Тилакоидная мембрана охватывает внутреннее пространство, называемое просветом тилакоида . Как показано на рисунке 5, стопка тилакоидов называется гранулом , а заполненное жидкостью пространство, окружающее гранум, называется стромой , или «ложе» (не путать со стомой или «ртом», отверстием на эпидермис листа).
Практический вопрос
Рис. 5. Фотосинтез происходит в хлоропластах, которые имеют внешнюю и внутреннюю мембраны. Стеки тилакоидов, называемые грана, образуют третий мембранный слой.
В жаркий и сухой день растения закрывают устьица для экономии воды. Какое влияние это окажет на фотосинтез?
Показать ответ
Уровень углекислого газа (необходимого фотосинтетического субстрата) немедленно упадет. В результате скорость фотосинтеза будет подавлена.
Две части фотосинтеза
Фотосинтез проходит в две последовательные стадии: светозависимые реакции и светонезависимые реакции. В светозависимых реакциях энергия солнечного света поглощается хлорофиллом, и эта энергия преобразуется в запасенную химическую энергию. В светонезависимых реакциях химическая энергия, собранная во время светозависимых реакций, управляет сборкой молекул сахара из углекислого газа.Следовательно, хотя светонезависимые реакции не используют свет в качестве реагента, они требуют, чтобы продукты светозависимых реакций функционировали. Кроме того, светом активируются несколько ферментов светонезависимых реакций. Светозависимые реакции используют определенные молекулы для временного хранения энергии: они называются энергоносителями. Энергоносители, которые перемещают энергию из светозависимых реакций в светонезависимые, можно рассматривать как «полные», потому что они богаты энергией.После высвобождения энергии «пустые» энергоносители возвращаются к светозависимой реакции, чтобы получить больше энергии. На рисунке 6 показаны компоненты внутри хлоропласта, где происходят светозависимые и светонезависимые реакции.
Рис. 6. Фотосинтез проходит в две стадии: светозависимые реакции и цикл Кальвина. Светозависимые реакции, которые происходят в тилакоидной мембране, используют световую энергию для образования АТФ и НАДФН. Цикл Кальвина, который имеет место в строме, использует энергию, полученную из этих соединений, для производства GA3P из CO 2 .
Щелкните ссылку, чтобы узнать больше о фотосинтезе.
Фотосинтез в продуктовом магазине
Рис. 7. Пища, потребляемая людьми, возникает в результате фотосинтеза. (кредит: Associação Brasileira de Supermercados)
Основные продуктовые магазины в Соединенных Штатах разделены на отделы, например, молочные продукты, мясо, продукты, хлеб, крупы и т. Д. В каждом проходе (рис. 7) находятся сотни, если не тысячи, различных продуктов, которые покупатели могут покупать и потреблять.
Хотя существует большое разнообразие, каждый элемент связан с фотосинтезом.Мясо и молочные продукты связаны, потому что животных кормили растительной пищей. Хлеб, крупы и макаронные изделия в основном получают из крахмалистых зерен, которые являются семенами растений, зависящих от фотосинтеза. А как насчет десертов и напитков? Все эти продукты содержат сахар — сахароза — это растительный продукт, дисахарид, молекула углеводов, которая строится непосредственно в результате фотосинтеза. Более того, многие предметы не так очевидно получены из растений: например, бумажные изделия, как правило, являются растительными продуктами, а многие пластмассы (в большом количестве в качестве продуктов и упаковки) могут быть получены из водорослей или из нефти, окаменелых остатков фотосинтезирующих организмов.Практически все специи и ароматизаторы в ряду специй были произведены растением в виде листа, корня, коры, цветка, фрукта или стебля. В конечном итоге фотосинтез связан с каждым приемом пищи и каждой пищей, которую человек потребляет.
Спектры света
Как можно использовать свет для приготовления пищи? Когда человек включает лампу, электрическая энергия становится световой. Как и все другие формы кинетической энергии, свет может перемещаться, изменять форму и использоваться для работы. В случае фотосинтеза световая энергия преобразуется в химическую энергию, которую фотоавтотрофы используют для создания молекул углеводов.Однако автотрофы используют только несколько определенных компонентов солнечного света.
Что такое световая энергия?
Солнце испускает огромное количество электромагнитного излучения (солнечной энергии). Люди могут видеть только часть этой энергии, поэтому эта часть называется «видимым светом». Способ распространения солнечной энергии описывается как волны. Ученые могут определить количество энергии волны, измерив ее длину волны, расстояние между последовательными точками волны.Одиночная волна измеряется от двух последовательных точек, например, от гребня к гребню или от впадины к впадине (Рисунок 8).
Рис. 8. Длина волны одиночной волны — это расстояние между двумя последовательными точками схожего положения (два гребня или две впадины) вдоль волны.
Видимый свет представляет собой только один из многих типов электромагнитного излучения, испускаемого Солнцем и другими звездами. Ученые различают различные типы лучистой энергии Солнца в пределах электромагнитного спектра.Электромагнитный спектр — это диапазон всех возможных частот излучения (рисунок 9). Разница между длинами волн связана с количеством переносимой ими энергии.
Рис. 9. Солнце излучает энергию в виде электромагнитного излучения. Это излучение существует на разных длинах волн, каждая из которых имеет свою характерную энергию. Все электромагнитное излучение, включая видимый свет, характеризуется длиной волны.
Каждый тип электромагнитного излучения распространяется на определенной длине волны.Чем больше длина волны (или чем больше она появляется на диаграмме), тем меньше энергии переносится. Короткие, плотные волны несут наибольшую энергию. Это может показаться нелогичным, но представьте это как кусок тяжелой веревки. Человеку не нужно прилагать особых усилий, чтобы переместить веревку длинными широкими волнами. Чтобы веревка двигалась короткими и крутыми волнами, человеку нужно приложить значительно больше энергии.
Электромагнитный спектр (рис. 9) показывает несколько типов электромагнитного излучения, исходящего от солнца, включая рентгеновские лучи и ультрафиолетовые (УФ) лучи.Волны более высокой энергии могут проникать в ткани и повреждать клетки и ДНК, что объясняет, почему и рентгеновские лучи, и ультрафиолетовые лучи могут быть вредными для живых организмов.
Поглощение света
Энергия света запускает процесс фотосинтеза, когда пигменты поглощают свет. Органические пигменты, будь то сетчатка глаза человека или тилакоид хлоропластов, имеют узкий диапазон уровней энергии, которые они могут поглощать. Уровни энергии ниже, чем те, которые представлены красным светом, недостаточны для поднятия орбитального электрона в населенное возбужденное (квантовое) состояние.Уровни энергии выше, чем в синем свете, физически разрывают молекулы на части, что называется обесцвечиванием. Таким образом, пигменты сетчатки могут «видеть» (поглощать) свет от 700 до 400 нм, который поэтому называется видимым светом. По тем же причинам молекулы пигментов растений поглощают только свет в диапазоне длин волн от 700 до 400 нм; физиологи растений называют этот диапазон для растений фотосинтетически активным излучением.
Видимый свет, который люди воспринимают как белый свет, на самом деле существует в радуге цветов.Некоторые объекты, такие как призма или капля воды, рассеивают белый свет, открывая цвета человеческому глазу. Часть видимого света электромагнитного спектра представляет собой радугу цветов, при этом фиолетовый и синий имеют более короткие длины волн и, следовательно, более высокую энергию. На другом конце спектра, ближе к красному, длины волн длиннее и имеют меньшую энергию (рис. 10).
Рис. 10. Цвета видимого света не несут такое же количество энергии. Фиолетовый имеет самую короткую длину волны и поэтому несет больше всего энергии, тогда как красный имеет самую длинную длину волны и несет наименьшее количество энергии.(кредит: модификация работы НАСА)
Общие сведения о пигментах
Существуют различные виды пигментов, каждый из которых эволюционировал, чтобы поглощать только определенные длины волн (цвета) видимого света. Пигменты отражают или пропускают длины волн, которые они не могут поглотить, благодаря чему они имеют соответствующий цвет.
Хлорофиллы и каротиноиды — два основных класса фотосинтетических пигментов, обнаруженных в растениях и водорослях; каждый класс имеет несколько типов молекул пигмента. Существует пять основных хлорофиллов: a , b , c и d и родственная молекула, обнаруженная в прокариотах, называемая бактериохлорофиллом. Хлорофилл a и хлорофилл b обнаружены в хлоропластах высших растений и будут в центре внимания следующего обсуждения.
Каротиноиды, имеющие множество различных форм, представляют собой гораздо большую группу пигментов. Каротиноиды, содержащиеся во фруктах, такие как красный цвет томата (ликопин), желтый цвет семян кукурузы (зеаксантин) или апельсин апельсиновой корки (β-каротин), используются в качестве рекламы для привлечения распространителей семян.При фотосинтезе
каротиноиды действуют как фотосинтетические пигменты, которые являются очень эффективными молекулами для утилизации избыточной энергии. Когда лист подвергается воздействию полного солнца, для обработки огромного количества энергии требуются светозависимые реакции; если с этой энергией не обращаться должным образом, она может нанести значительный ущерб. Следовательно, многие каротиноиды находятся в тилакоидной мембране, поглощают избыточную энергию и безопасно рассеивают эту энергию в виде тепла.
Каждый тип пигмента можно идентифицировать по определенному спектру длин волн, который он поглощает из видимого света, то есть спектру поглощения .График на Рисунке 11 показывает спектры поглощения хлорофилла a , хлорофилла b и типа каротиноидного пигмента, называемого β-каротином (который поглощает синий и зеленый свет). Обратите внимание, как у каждого пигмента есть свой набор пиков и впадин, раскрывающих очень специфический образец поглощения. Хлорофилл a поглощает длины волн с любого конца видимого спектра (синий и красный), но не зеленого цвета. Поскольку зеленый цвет отражается или передается, хлорофилл кажется зеленым.Каротиноиды поглощают в коротковолновой синей области и отражают более длинные волны желтого, красного и оранжевого цветов.
Рис. 11. (a) Хлорофилл a, (b) хлорофилл b и (c) β-каротин — это гидрофобные органические пигменты, обнаруженные в тилакоидной мембране. Хлорофиллы a и b, которые идентичны, за исключением части, указанной в красном поле, отвечают за зеленый цвет листьев. β-каротин отвечает за оранжевый цвет моркови. Каждый пигмент имеет (г) уникальный спектр поглощения.
Рис. 12. Растения, которые обычно растут в тени, адаптировались к низким уровням света за счет изменения относительной концентрации пигментов хлорофилла. (кредит: Джейсон Холлингер)
Многие фотосинтезирующие организмы имеют смесь пигментов; с их помощью организм может поглощать энергию более широкого диапазона длин волн. Не все фотосинтезирующие организмы имеют полный доступ к солнечному свету. Некоторые организмы растут под водой, где интенсивность и качество света уменьшаются и меняются с глубиной.Другие организмы растут в конкуренции за свет. Растения на полу тропического леса должны поглощать любой проникающий свет, потому что более высокие деревья поглощают большую часть солнечного света и рассеивают оставшееся солнечное излучение (рис. 12).
При изучении фотосинтезирующих организмов ученые могут определять типы присутствующих пигментов, генерируя спектры поглощения. Прибор, называемый спектрофотометром , может различать световые волны с длиной волны, которые может поглощать вещество.Спектрофотометры измеряют проходящий свет и вычисляют по нему поглощение. Извлекая пигменты из листьев и помещая эти образцы в спектрофотометр, ученые могут определить, какие длины волн света может поглощать организм. Дополнительные методы идентификации растительных пигментов включают различные типы хроматографии, которые разделяют пигменты по их относительному сродству к твердой и подвижной фазам.
Светозависимые реакции
Общая функция светозависимых реакций состоит в преобразовании солнечной энергии в химическую энергию в форме НАДФН и АТФ.Эта химическая энергия поддерживает светонезависимые реакции и способствует сборке молекул сахара. Светозависимые реакции изображены на рисунке 13. Белковые комплексы и молекулы пигмента работают вместе, чтобы производить НАДФН и АТФ.
Рис. 13. Фотосистема состоит из светособирающего комплекса и реакционного центра. Пигменты светособирающего комплекса передают световую энергию двум особым молекулам хлорофилла а в реакционном центре. Свет возбуждает электрон от пары хлорофилла А, который переходит к первичному акцептору электронов.Затем необходимо заменить возбужденный электрон. В (а) фотосистеме II электрон возникает в результате расщепления воды, которая выделяет кислород в качестве побочного продукта. В (б) фотосистеме I электрон исходит из цепи переноса электронов хлоропласта, обсуждаемой ниже.
Фактический этап преобразования световой энергии в химическую энергию происходит в мультибелковом комплексе, называемом фотосистемой , два типа которой встроены в тилакоидную мембрану, фотосистема II (PSII) и фотосистема I (PSI). (Рисунок 14).Эти два комплекса различаются в зависимости от того, что они окисляют (то есть источник поступления низкоэнергетических электронов) и что они восстанавливают (место, куда они доставляют свои заряженные электроны).
Обе фотосистемы имеют одинаковую базовую структуру; ряд антенных белков, с которыми связаны молекулы хлорофилла, окружают реакционный центр, в котором происходит фотохимия. Каждая фотосистема обслуживается светособирающим комплексом, который передает энергию солнечного света в реакционный центр; он состоит из множественных антенных белков, которые содержат смесь 300–400 молекул хлорофилла a и b , а также другие пигменты, такие как каротиноиды.Поглощение одного фотона или определенного количества или «пакета» света любым из хлорофиллов переводит эту молекулу в возбужденное состояние. Короче говоря, световая энергия теперь улавливается биологическими молекулами, но еще не хранится в какой-либо полезной форме. Энергия передается от хлорофилла к хлорофиллу до тех пор, пока в конечном итоге (примерно через одну миллионную секунды) не будет доставлена в реакционный центр. До этого момента между молекулами передавалась только энергия, а не электроны.
Практический вопрос
Рис. 14. Реакционный центр фотосистемы II (ФСII) и фотосистемы I (ФСI).
В реакционном центре фотосистемы II (ФСII) энергия солнечного света используется для извлечения электронов из воды. Электроны перемещаются по цепи переноса электронов хлоропласта в фотосистему I (PSI), которая восстанавливает NADP + до NADPH. Цепь переноса электронов перемещает протоны через тилакоидную мембрану в просвет. В то же время расщепление воды добавляет протоны к просвету, а уменьшение НАДФН удаляет протоны из стромы.Конечный результат — низкий pH в просвете тилакоидов и высокий pH в строме. АТФ-синтаза использует этот электрохимический градиент для производства АТФ. Что является исходным источником электронов для цепи переноса электронов хлоропласта?
- вода
- кислород
- углекислый газ
- НАДФН
Показать ответ
Вода — исходный источник.
Реакционный центр содержит пару молекул хлорофилла и с особым свойством.Эти два хлорофилла могут окисляться при возбуждении; они действительно могут отдать электрон в процессе, называемом фотоакт . Именно на этой стадии в реакционном центре световая энергия преобразуется в возбужденный электрон. Все последующие шаги включают перенос этого электрона на носитель энергии НАДФН для доставки в цикл Кальвина, где электрон откладывается на углероде для длительного хранения в форме углевода. PSII и PSI являются двумя основными компонентами фотосинтетической цепи переноса электронов , которая также включает цитохромный комплекс .Цитохромный комплекс, фермент, состоящий из двух белковых комплексов, переносит электроны от молекулы-носителя пластохинона (Pq) к белку пластоцианину (Pc), тем самым обеспечивая перенос протонов через тилакоидную мембрану и перенос электронов от ФСII к PSI.
Реакционный центр PSII (называемый P680 ) доставляет свои высокоэнергетические электроны по одному к первичному акцептору электронов и через цепь переноса электронов (Pq в комплекс цитохрома и пластоцианин) в PSI.Пропавший электрон P680 заменяется извлечением электрона с низкой энергией из воды; таким образом, вода разделяется, и ФСII восстанавливается после каждого фотоакта. При расщеплении одной молекулы H 2 O выделяются два электрона, два атома водорода и один атом кислорода. Для образования одной молекулы двухатомного газа O 2 требуется разделение двух молекул. Около 10 процентов кислорода используется митохондриями листа для поддержки окислительного фосфорилирования. Остаток улетучивается в атмосферу, где используется аэробными организмами для поддержки дыхания.
Когда электроны движутся через белки, расположенные между PSII и PSI, они теряют энергию. Эта энергия используется для перемещения атомов водорода со стромальной стороны мембраны в просвет тилакоида. Эти атомы водорода, а также те, которые образуются при расщеплении воды, накапливаются в просвете тилакоида и будут использоваться для синтеза АТФ на более позднем этапе. Поскольку электроны потеряли энергию до своего прибытия в PSI, они должны быть повторно активированы PSI, следовательно, другой фотон поглощается антенной PSI.Эта энергия передается в реакционный центр PSI (называемый P700 ). P700 окисляется и отправляет электрон высокой энергии на NADP + с образованием NADPH. Таким образом, PSII захватывает энергию для создания протонных градиентов для образования АТФ, а PSI захватывает энергию для восстановления NADP + в NADPH. Две фотосистемы работают согласованно, отчасти, чтобы гарантировать, что производство НАДФН будет примерно равно производству АТФ. Существуют и другие механизмы для точной настройки этого соотношения, чтобы точно соответствовать постоянно меняющимся потребностям хлоропласта в энергии.
Создание носителя энергии: ATP
Как и в межмембранном пространстве митохондрий во время клеточного дыхания, накопление ионов водорода внутри просвета тилакоидов создает градиент концентрации. Пассивная диффузия ионов водорода от высокой концентрации (в просвете тилакоида) до низкой концентрации (в строме) используется для создания АТФ, как и в цепи переноса электронов клеточного дыхания. Ионы накапливают энергию из-за диффузии и из-за того, что все они имеют одинаковый электрический заряд, отталкивая друг друга.
Чтобы высвободить эту энергию, ионы водорода устремляются через любое отверстие, как струя воды через отверстие в дамбе. В тилакоиде это отверстие представляет собой проход через специализированный белковый канал, называемый АТФ-синтазой. Энергия, выделяемая потоком ионов водорода, позволяет АТФ-синтазе присоединять третью фосфатную группу к АДФ, которая образует молекулу АТФ (рис. 14). Поток ионов водорода через АТФ-синтазу называется хемиосмосом, потому что ионы перемещаются из области с высокой концентрацией в область с низкой концентрацией через полупроницаемую структуру.
Посетите этот сайт и просмотрите анимацию, чтобы увидеть процесс фотосинтеза в листе.
Светонезависимые реакции
После того, как энергия солнца преобразуется в химическую энергию и временно сохраняется в молекулах АТФ и НАДФН, у клетки появляется топливо, необходимое для создания молекул углеводов для длительного хранения энергии. Продукты светозависимых реакций, АТФ и НАДФН, имеют продолжительность жизни в диапазоне миллионных долей секунды, тогда как продукты светонезависимых реакций (углеводы и другие формы восстановленного углерода) могут сохраняться в течение сотен миллионов лет.Созданные молекулы углеводов будут иметь основу из атомов углерода. Откуда берется углерод? Он происходит из углекислого газа, газа, который является побочным продуктом дыхания микробов, грибов, растений и животных.
У растений углекислый газ (CO 2 ) попадает в листья через устьица, где он диффундирует на короткие расстояния через межклеточные пространства, пока не достигнет клеток мезофилла. Попадая в клетки мезофилла, CO 2 диффундирует в строму хлоропласта — место светонезависимых реакций фотосинтеза.Эти реакции на самом деле имеют несколько названий. Другой термин, цикл Кальвина , назван в честь человека, который его открыл, и потому, что эти реакции функционируют как цикл. Другие называют это циклом Кальвина-Бенсона, чтобы включить имя другого ученого, участвовавшего в его открытии. Наиболее устаревшее название — темные реакции, потому что свет напрямую не требуется (рис. 15). Однако термин «темная реакция» может вводить в заблуждение, поскольку он неверно подразумевает, что реакция происходит только ночью или не зависит от света, поэтому большинство ученых и инструкторов больше не используют его.
Рис. 15. Световые реакции используют энергию солнца для образования химических связей, АТФ и НАДФН. Эти несущие энергию молекулы образуются в строме, где происходит фиксация углерода.
Независимые от света реакции цикла Кальвина можно разделить на три основных этапа: фиксация, восстановление и регенерация.
Этап 1: Фиксация
В строме, помимо CO 2 , присутствуют два других компонента, инициирующих светонезависимые реакции: фермент, называемый рибулозобисфосфаткарбоксилазой (RuBisCO), и три молекулы рибулозобисфосфата (RuBP), как показано на рисунке 16.RuBP имеет пять атомов углерода, окруженных двумя фосфатами.
Практический вопрос
Рисунок 16. Цикл Кальвина состоит из трех этапов.
На стадии 1 фермент RuBisCO включает диоксид углерода в органическую молекулу 3-PGA. На стадии 2 органическая молекула восстанавливается с помощью электронов, поставляемых НАДФН. На стадии 3 RuBP, молекула, которая запускает цикл, регенерируется, так что цикл может продолжаться. Только одна молекула углекислого газа включается за один раз, поэтому цикл должен быть завершен три раза, чтобы произвести одну молекулу GA3P с тремя атомами углерода, и шесть раз, чтобы произвести молекулу глюкозы с шестью атомами углерода.
Какое из следующих утверждений верно?
- При фотосинтезе реагентами являются кислород, углекислый газ, АТФ и НАДФН. GA3P и вода являются продуктами.
- Реагентами фотосинтеза являются хлорофилл, вода и углекислый газ. GA3P и кислород являются продуктами.
- При фотосинтезе реагентами являются вода, углекислый газ, АТФ и НАДФН. RuBP и кислород — это продукты.
- Реагентами фотосинтеза являются вода и углекислый газ. GA3P и кислород являются продуктами.
Показать ответ
Ответ d верный.
RuBisCO катализирует реакцию между CO 2 и RuBP. Для каждой молекулы CO 2 , которая реагирует с одним RuBP, образуются две молекулы другого соединения (3-PGA). PGA имеет три атома углерода и один фосфат. Каждый виток цикла включает только один RuBP и один диоксид углерода и образует две молекулы 3-PGA. Число атомов углерода остается тем же, поскольку атомы перемещаются, чтобы образовывать новые связи во время реакций (3 атома из 3CO 2 + 15 атомов из 3RuBP = 18 атомов в 3 атомах 3-PGA).Этот процесс называется фиксация углерода , потому что CO 2 «фиксируется» из неорганической формы в органические молекулы.
Этап 2: Редукция
АТФ и НАДФН используются для преобразования шести молекул 3-PGA в шесть молекул химического вещества, называемого глицеральдегид-3-фосфатом (G3P). Это реакция восстановления, потому что она включает в себя усиление электронов 3-PGA. Напомним, что уменьшение — это усиление электрона атомом или молекулой. Используются шесть молекул как АТФ, так и НАДФН.Для АТФ энергия высвобождается с потерей концевого атома фосфата, превращая его в АДФ; для НАДФН теряется энергия и атом водорода, превращая его в НАДФ + . Обе эти молекулы возвращаются к ближайшим светозависимым реакциям для повторного использования и восстановления энергии.
Этап 3: Регенерация
Интересно, что в этот момент только одна из молекул G3P покидает цикл Кальвина и отправляется в цитоплазму, чтобы способствовать образованию других соединений, необходимых для растения.Поскольку G3P, экспортированный из хлоропласта, имеет три атома углерода, требуется три «витка» цикла Кальвина, чтобы зафиксировать достаточно чистого углерода для экспорта одного G3P. Но каждый ход дает два G3P, таким образом, три хода составляют шесть G3P. Одна экспортируется, а остальные пять молекул G3P остаются в цикле и используются для регенерации RuBP, что позволяет системе подготовиться к фиксации большего количества CO 2 . В этих реакциях регенерации используются еще три молекулы АТФ.
Эта ссылка ведет к анимации цикла Кальвина.Щелкните этап 1, этап 2, а затем этап 3, чтобы увидеть, как G3P и ATP регенерируют с образованием RuBP.
Эволюция фотосинтеза
Рис. 17. Суровые условия пустыни побудили такие растения, как эти кактусы, развить вариации светонезависимых реакций фотосинтеза. Эти изменения повышают эффективность использования воды, помогая экономить воду и энергию. (кредит: Петр Войтковский)
В ходе эволюции фотосинтеза произошел значительный сдвиг от бактериального типа фотосинтеза, который включает только одну фотосистему и обычно является аноксигенным (не генерирует кислород), к современному оксигеническому (действительно генерирующему кислород) фотосинтезу с использованием двух фотосистем.Этот современный кислородный фотосинтез используется многими организмами — от гигантских тропических листьев в тропических лесах до крошечных цианобактериальных клеток — и процесс и компоненты этого фотосинтеза остаются в основном такими же. Фотосистемы поглощают свет и используют цепи переноса электронов для преобразования энергии в химическую энергию АТФ и НАДН. Последующие светонезависимые реакции собирают молекулы углеводов с этой энергией.
Фотосинтез у пустынных растений привел к развитию адаптаций, которые экономят воду.В суровой и сухой жаре нужно использовать каждую каплю воды, чтобы выжить. Поскольку устьица должны открываться, чтобы обеспечить поглощение CO 2 , вода уходит из листа во время активного фотосинтеза. Пустынные растения разработали способы экономии воды и работы в суровых условиях. Более эффективное использование CO 2 позволяет растениям адаптироваться к жизни с меньшим количеством воды. Некоторые растения, такие как кактусы (рис. 17), могут подготавливать материалы для фотосинтеза в ночное время за счет временного процесса фиксации / хранения углерода, поскольку открытие устьиц в это время сохраняет воду из-за более низких температур.Кроме того, кактусы развили способность осуществлять низкий уровень фотосинтеза, вообще не открывая устьиц, — механизм, позволяющий справляться с чрезвычайно засушливыми периодами.
Теперь, когда мы узнали о различных аспектах фотосинтеза, давайте соберем все вместе. Это видео проведет вас через процесс фотосинтеза в целом:
Вкратце: обзор фотосинтеза
Процесс фотосинтеза изменил жизнь на Земле. Используя энергию солнца, фотосинтез эволюционировал, чтобы позволить живым существам получить доступ к огромному количеству энергии.Благодаря фотосинтезу живые существа получили доступ к достаточному количеству энергии, что позволило им построить новые структуры и достичь очевидного сегодня биоразнообразия.
Только некоторые организмы, называемые фотоавтотрофами, могут осуществлять фотосинтез; они требуют наличия хлорофилла, специального пигмента, который поглощает определенные части видимого спектра и может улавливать энергию солнечного света. Фотосинтез использует углекислый газ и воду для сборки молекул углеводов и выделения кислорода в виде отходов в атмосферу.У эукариотических автотрофов, таких как растения и водоросли, есть органеллы, называемые хлоропластами, в которых происходит фотосинтез и накапливается крахмал. У прокариот, таких как цианобактерии, этот процесс менее локализован и происходит внутри складчатых мембран, выступов плазматической мембраны и в цитоплазме.
Пигменты первой части фотосинтеза, светозависимые реакции, поглощают энергию солнечного света. Фотон ударяет антенные пигменты фотосистемы II, чтобы инициировать фотосинтез.Энергия передается в реакционный центр, содержащий хлорофилл и , к цепи переноса электронов, которая закачивает ионы водорода внутрь тилакоида. Это действие создает высокую концентрацию ионов. Ионы проходят через АТФ-синтазу через хемиосмос с образованием молекул АТФ, которые используются для образования молекул сахара на второй стадии фотосинтеза. Фотосистема I поглощает второй фотон, что приводит к образованию молекулы НАДФН, другого носителя энергии и снижения энергии для светонезависимых реакций.
Проверьте свое понимание
Ответьте на вопросы ниже, чтобы увидеть, насколько хорошо вы понимаете темы, затронутые в предыдущем разделе. В этой короткой викторине , а не засчитываются в вашу оценку в классе, и вы можете пересдавать ее неограниченное количество раз.
Используйте этот тест, чтобы проверить свое понимание и решить, следует ли (1) изучить предыдущий раздел дальше или (2) перейти к следующему разделу.
Модуляция активности ферментов при фотосинтезе светом / темнотой в JSTOR
Abstract
У фотосинтезирующих видов, от цианобактерий до высших растений, многие ферменты модулируются светом.Большинство известных ферментов с модуляцией света являются хлоропластами. Было предложено четыре механизма модуляции. Одной из функций модуляции света, вероятно, является автокаталитическое накопление промежуточных продуктов восстановительного пентозофосфатного цикла во время индукционного периода фотосинтетической фиксации CO2.
Journal Information
Изданный Американским институтом биологических наук (AIBS), BioScience представляет читателям своевременные и авторитетные обзоры текущих исследований в области биологии, сопровождаемые эссе и дискуссионными разделами по образованию, государственной политике, истории и концептуальным основам биологических исследований. науки.Рецензируемый, часто цитируемый ежемесячный журнал с содержанием, написанным и отредактированным для доступа исследователей, преподавателей и студентов, BioScience включает статьи о результатах и методах исследований, достижениях в области биологического образования, профессионально написанные тематические статьи о последних рубежах в биологии. , обсуждение профессиональных вопросов, обзоры книг, новости об AIBS, политическая колонка (Washington Watch) и образовательная колонка (Eye on Education). Круглые столы, форумы и статьи о точках зрения представляют точки зрения лидеров мнений и приглашают к дальнейшим комментариям.BioScience входит в число лучших журналов в своей категории ISI (биология) как по импакт-фактору, так и по периодам полураспада цитирования.
Информация об издателе
Oxford University Press — это отделение Оксфордского университета. Издание во всем мире способствует достижению цели университета в области исследований, стипендий и образования. OUP — крупнейшая в мире университетская пресса с самым широким глобальным присутствием. В настоящее время он издает более 6000 новых публикаций в год, имеет офисы примерно в пятидесяти странах и насчитывает более 5500 сотрудников по всему миру.Он стал известен миллионам людей благодаря разнообразной издательской программе, которая включает научные работы по всем академическим дисциплинам, библии, музыку, школьные и университетские учебники, книги по бизнесу, словари и справочники, а также академические журналы.
Фотосинтез — биохимия
Если вы считаете, что контент, доступный через Веб-сайт (как определено в наших Условиях обслуживания), нарушает одно
или другие ваши авторские права, сообщите нам, отправив письменное уведомление («Уведомление о нарушении»), содержащее
в
информацию, описанную ниже, назначенному ниже агенту.Если репетиторы университета предпримут действия в ответ на
ан
Уведомление о нарушении, оно предпримет добросовестную попытку связаться со стороной, которая предоставила такой контент
средствами самого последнего адреса электронной почты, если таковой имеется, предоставленного такой стороной Varsity Tutors.
Ваше Уведомление о нарушении прав может быть отправлено стороне, предоставившей доступ к контенту, или третьим лицам, таким как
в виде
ChillingEffects.org.
Обратите внимание, что вы будете нести ответственность за ущерб (включая расходы и гонорары адвокатов), если вы существенно
искажать информацию о том, что продукт или действие нарушает ваши авторские права.Таким образом, если вы не уверены, что контент находится
на Веб-сайте или по ссылке с него нарушает ваши авторские права, вам следует сначала обратиться к юристу.
Чтобы отправить уведомление, выполните следующие действия:
Вы должны включить следующее:
Физическая или электронная подпись правообладателя или лица, уполномоченного действовать от их имени;
Идентификация авторских прав, которые, как утверждается, были нарушены;
Описание характера и точного местонахождения контента, который, по вашему мнению, нарушает ваши авторские права, в \
достаточно подробностей, чтобы позволить репетиторам университетских школ найти и точно идентифицировать этот контент; например нам требуется
а
ссылка на конкретный вопрос (а не только на название вопроса), который содержит содержание и описание
к какой конкретной части вопроса — изображению, ссылке, тексту и т. д. — относится ваша жалоба;
Ваше имя, адрес, номер телефона и адрес электронной почты; а также
Ваше заявление: (а) вы добросовестно полагаете, что использование контента, который, по вашему мнению, нарушает
ваши авторские права не разрешены законом, владельцем авторских прав или его агентом; (б) что все
информация, содержащаяся в вашем Уведомлении о нарушении, является точной, и (c) под страхом наказания за лжесвидетельство, что вы
либо владелец авторских прав, либо лицо, уполномоченное действовать от их имени.
Отправьте жалобу нашему уполномоченному агенту по адресу:
Чарльз Кон
Varsity Tutors LLC
101 S. Hanley Rd, Suite 300
St. Louis, MO 63105
Или заполните форму ниже:
Активность фермента
при фотосинтезе | Education
Фотосинтез — это сложная серия реакций, которые растения и водоросли используют для преобразования световой энергии, воды и углекислого газа в глюкозу.Фотосинтез происходит в два этапа: световые реакции и темные реакции. В световых реакциях пигментный хлорофилл поглощает световую энергию и использует ее для создания высокоэнергетических молекул АТФ и НАДФН. В темновых реакциях АТФ и НАДФН используются для фиксации углерода в цикле Кальвина.
Световые реакции
В хлоропластах растительных клеток есть ряд ферментов и других белков, называемых фотосистемами I и II. Некоторые из этих белков содержат молекулы хлорофилла, и когда на хлорофилл попадает фотография света, электрон получает энергию, и молекулы воды связываются.Фермент фотосистемы II расщепляет воду на водород и кислород. Затем заряженный электрон проходит через другой фермент, называемый комплексом цитохрома b6-f, который, в свою очередь, перекачивает положительно заряженные молекулы водорода через мембрану для создания запаса потенциальной энергии. Фермент под названием АТФ-синтаза использует эту потенциальную энергию для синтеза молекулы под названием АТФ, которая является наиболее распространенной энергетической валютой для клеток и которая используется в темных реакциях фотосинтеза.
Хлоропласты содержат второй механизм захвата света, называемый фотосистемой I, который действует аналогично фотосистеме II, за исключением того, что конечным результатом является молекула под названием НАДФН вместо АТФ.Фотосистема I связана с фотосистемой II небольшим белком, называемым пластоцианин, который отдает электроны из фотосистемы II фотосистеме I.
Цикл Кальвина
В цикле Кальвина АТФ и НАДФН используются для уменьшения или фиксации углекислого газа. производят глюкозу. Эта реакция связывания углерода катализируется ферментом под названием RUBISCO, белком в большом количестве в растениях и, возможно, самым распространенным белком на Земле. Одна за другой три молекулы углекислого газа поглощаются RUBISCO и двумя другими ферментами для производства одной молекулы трехуглеродного сахара, называемого глицеральдегид-3-фосфатом.Две молекулы глицеральдегид-3-фосфата в конечном итоге объединяются в одну молекулу глюкозы.
C3 и C4 Растения
Фиксация углерода, приводящая к трехуглеродному сахару, известна как фотосинтез C3, и большинство растений используют этот вид фотосинтеза. Он хорошо работает в прохладных и влажных условиях, когда растения могут сохранять открытыми поры, которые пропускают углекислый газ, называемые устьицами, в течение дня.
Для многих летних однолетних растений условия слишком жаркие, чтобы устьица оставались открытыми в течение дня, поэтому эти растения адаптировались, развивая фотосинтез С4.Фотосинтез C4 назван так потому, что фиксация углекислого газа ферментом, называемым PEP-карбоксилазой, приводит к образованию четырехуглеродного сахара. Полученный таким образом углерод затем передается в RUBISCO и входит в цикл Кальвина.
CAM-растения
Третий тип фотосинтеза известен как CAM-дыхание. Суккуленты и некоторые орхидеи выполняют САМ-дыхание. Эти растения открывают устьица ночью и накапливают углерод из углекислого газа в виде кислоты толстолистых. В дневное время, когда энергия для синтеза АТФ и НАДФН поступает от света, углерод высвобождается из крассулоидной кислоты и передается в RUBISCO, чтобы начать цикл Кальвина.
Светонезависимые реакции — обзор
15.1 Введение
Физиологическое состояние нескольких компонентов фотосистемы II (ФСII), компонентов цепи переноса электронов и взаимодействие светозависимых и светонезависимых биохимических реакций можно оценить с помощью анализа. кривых индукции флуоресценции хлорофилла (ChlF) (Govindjee, 1995; DeEll and Toivonen, 2003; Papageorgiou and Govindjee, 2004; Strasser et al., 2004; Kalaji et al., 2011; Suggett et al., 2011). Тесное сотрудничество и точная регулировка скорости этих реакций необходимы для того, чтобы весь процесс фотосинтеза протекал с высокой эффективностью (Murkowski, 2002; Van der Tol et al., 2009). Измерения ChlF позволяют распознавать изменения в общем биоэнергетическом состоянии фотосинтетического аппарата. Более того, такие измерения прямо или косвенно относятся ко всем стадиям светозависимых фотосинтетических реакций, включая расщепление воды, перенос электронов и формирование градиента pH через тилакоидную мембрану и синтез АТФ (Kalaji et al., 2012).
В настоящее время измерения ChlF являются популярным методом оценки воздействия стрессовых факторов на фотосинтез. Они представляют собой простой, недеструктивный, недорогой и быстрый инструмент для анализа светозависимых фотосинтетических реакций и для косвенной оценки содержания хлорофилла в одном и том же образце ткани. (Базовую информацию о связи ChlF с фотосинтезом см. В обзорах Papageorgiou and Govindjee, 2011, и Stirbet and Govindjee (2011, 2012). Улучшенные методы измерения ChlF позволяют изучать процесс фотосинтеза на различных функциональных уровнях (e .g., светособирающие комплексы, первичные световые реакции, электронная транспортная цепь, светонезависимые реакции в тилакоидной строме и даже медленные регуляторные процессы). Ученые из разных областей, таких как физиология растений, биотехнология, лесоводство, экофизиология, и даже селекционеры и фермеры, ценят технические преимущества ChlF и с его помощью изучают структуру и функции фотосинтетического аппарата.
Значение параметров, полученных из анализа кинетики ChlF, можно использовать для исследования прямых фотосинтетических реакций на неблагоприятные условия окружающей среды и для косвенной оценки стрессовых воздействий на растения (Kuckenberg et al., 2009). Тщательно выбранные методы ChlF позволяют определять реакции на различные компоненты окружающей среды, такие как почва (дефицит питательных веществ, засоление, содержание тяжелых металлов), атмосфера (качество и интенсивность PAR, высокие и низкие температуры, содержание озона) и гербициды ( Fracheboud, Leipner, 2003; Dai et al., 2009; Buonasera et al., 2011). Исследования могут проводиться на одном растении (включая трансгенные и культивируемые in vitro) или на целых экосистемах для оценки потенциальной продуктивности и устойчивости к отдельным или группам стрессовых факторов (Bolhár-Nordenkampf and Öquist, 1993; Schreiber et al., 1994; Крупа и Башинский, 1995; Максвелл и Джонсон, 2000; Штрассер и др., 2000; Мурковский, 2002). Используя измерения ChlF, есть возможность обнаружить влияние стрессовых факторов до того, как станут заметны морфологические симптомы (например, увядание, некроз и хлороз) или даже до того, как станут очевидными изменения в содержании хлорофилла (Деви и Прасад, 1996; Рощина и Мельникова) , 1996; Kalaji and Pietkiewicz, 2004; Strasser et al., 2007; Kalaji and Guo, 2008; Tsimilli-Michael and Strasser, 2008; Kuckenberg et al., 2009).
ChlF потенциально имеет широкое применение, поскольку хлорофилл a присутствует во всех организмах, способных к кислородному фотосинтезу (эмбриофиты, водоросли, лишайники и цианобактерии). Методы исследований, основанные на измерениях ChlF, эффективно используются в ряде областей исследований, таких как физиология растений и защита растений (Merz et al., 1996), биоэнергетика (Wrochna et al., 2007), сельское хозяйство (Murkowski, 2002), садоводство. (Flexas et al., 2002; Burzyński, urek, 2007), лесное хозяйство (Mohammed et al., 1995; Percival et al., 2006), биология растений (Prasil et al., 1992), биотехнология (Takahashi et al., 2007), селекция растений (Kalaji and Pietkiewicz, 2004; Rykaczewska et al., 2004; Kalaji and Guo, 2008). ), экологии (Roger and Weiss, 2003; Chernev et al., 2006), складирования овощей и фруктов (Nedbal et al., 2001), а также пищевых технологий и переработки (Kuckenberg et al., 2008; Qiu et al., 2013). Измерения ChlF также полезны для оценки качества фруктов, овощей и цветов и для определения наилучшего времени для их продажи (Merz et al., 1996; Nedbal et al., 2001). Их также можно использовать для определения зрелости семян (Jalink et al., 1998), для оценки качества воды (Romanowska-Duda et al., 2005) и для оценки аллопатического воздействия вторичных метаболитов на рост и развитие растений (Devi and Прасад, 1996; Рощина, Мельникова, 1996).
Методы измерения, основанные на ChlF, являются одними из наиболее важных инструментов, используемых в настоящее время в программах селекции растений из-за их способности определять физиологическое состояние растений, обеспечивая понимание роста и урожайности растений в естественных стрессовых условиях окружающей среды (Kalaji and Guo, 2008 ).Некоторые параметры ChlF рекомендуются в качестве надежных биомаркеров для различных вариантов выбора толерантности к определенным группам гербицидов (например, гербицидам мочевины, триазина и диазина) (Devine et al., 1992; Dewez et al., 2008). Одним из наиболее многообещающих методов автоматической идентификации видов растений являются нейронные сети, в которых кривая индукции ChlF является одним из основных компонентов (Codrea et al., 2004). Таким образом, как видно из предыдущего, было разработано множество методик и приложений ChlF.Все они способствуют расширению знаний о фотосинтезе. В этой главе мы сосредоточимся в основном на анализе быстрой флуоресценции (до 1 с), вызванной непрерывным освещением и полученной с помощью JIP-теста, надежной математической модели, предложенной Штрассером и др. (2004); он содержит подробную информацию о состоянии и функции реакционных центров ФСII, антенны, а также соединений донорной и акцепторной сторон ФСII. Однако основная цель этой работы — описать влияние стрессовых факторов (стрессоров) на фотохимические процессы, отражающееся в изменениях кинетики быстрой ChlF и связанных с ней биофизических параметров.
Темные реакции фотосинтеза: цикл Кальвина-Бенсона — стенограмма видео и урока
Цикл Кальвина
Ключ к созданию сахаров — это фиксация углерода , когда неорганический углерод фиксируется в органическом углероде, таком как сахара. В цикле Кальвина углекислый газ превращается в органическую молекулу с помощью фермента под названием RuBisCO . Здесь углекислый газ, который мы выдыхаем как отходы, используется растениями, выполняющими фотосинтез.Давайте выделим некоторые этапы этого цикла Кальвина.
RuBisCO объединяет три молекулы углекислого газа с тремя 5-углеродными молекулами, каждая из которых имеет два фосфата, называемых RuBP . Создаются шесть 3-углеродных молекул, каждая из которых содержит один фосфат. Затем используются шесть молекул АТФ, отдавая каждой молекуле еще один фосфат. Затем шесть электронных носителей НАДФН из легких реакций переносят электроны в этот цикл Кальвина. Они окисляются до НАДФ +, отдавая электроны молекулам углерода, которые затем восстанавливаются.В результате образуются шесть суперэнергетических 3-углеродных молекул, называемых глицеральдегид-3-фосфатом, или G3P . Это делает молекулу из 3 атомов углерода с одной фосфатной группой. Одна из этих счастливых молекул G3P используется для создания глюкозы или других молекул углерода при выходе из цикла, в то время как остальные пять молекул перестраиваются с помощью трех АТФ, которые воссоздают три молекулы RuBP, которые могут снова начать цикл. Это регенерация RuBP, позволяющая продолжить цикл Кальвина.
Молекула G3P, выходящая из цикла, является наиболее важной.Две молекулы G3P могут создать одну молекулу глюкозы. Эта молекула глюкозы является строительным блоком для создания всего остального в растении и питает все остальные организмы, питающиеся растениями, такие как мы.
Когда фотодыхание заставляет цикл Кальвина повернуть не в ту сторону
Давайте вернемся к той первой стадии цикла Кальвина. На этом этапе три молекулы диоксида углерода присоединяются к трем молекулам из 5 атомов углерода, каждая из которых содержит два фосфата. Но вместо того, чтобы просто собрать нужное количество углекислого газа и превратить его в 5-углеродные молекулы, аккуратно продолжая цикл Кальвина, иногда фермент RuBisCO действует так, как будто у него есть собственный разум.Вместо этого он поглощает кислород, инициируя процесс, называемый фотодыханием .
Фотодыхание — это побочная реакция, которая иногда возникает во время цикла Кальвина. Он сводит на нет то хорошее, что обычно делает фотосинтез, потому что заставляет углерод, который уже был закреплен в органических соединениях, превращаться обратно в углекислый газ, откуда он пришел. Это снижает количество синтезируемой глюкозы.
Все проблемы начинаются с RuBisCO, который является непостоянным ферментом, потому что он может связываться как с углекислым газом, так и с молекулярным кислородом.Но что определяет, какой из них он выберет? Факторы, которые определяют, кто будет на свидании RuBisCO на вечер, — это относительная концентрация углекислого газа по отношению к кислороду и температура окружающей среды. Когда поры листьев или устьица растения открыты, кислород и водяной пар рассеиваются наружу. атмосфера. Однако углекислый газ попадает в растение. Итак, когда устьица открыты, фотореспиратона гораздо меньше, так как RuBisCO связывается с углекислым газом. Однако, когда устьица закрыты, кислород может накапливаться внутри листа растения, и тогда более вероятно возникновение фотодыхания.Непостоянный RuBisCO также имеет более высокое сродство к кислороду при повышении температуры окружающей среды, поэтому фотодыхание более вероятно в более жарких условиях.
Краткое содержание урока
В этом уроке мы узнали, как темная реакция или цикл Кальвина используют углекислый газ из атмосферы, а также АТФ и НАДФН из световых реакций для производства сахара. Помните, что в световых реакциях также использовалась вода и выделялся кислород. Три молекулы CO2 используются для фиксации углерода ферментом RuBisCO.За один раз используется один углекислый газ, что приводит к этапам, которые фиксируют, перестраивают и активируют молекулы углерода, в конечном итоге образуя одну 3-углеродную молекулу, называемую G3P , или глицеральдегид-3-фосфат, которая высвобождается.